

Circular permutation prediction reveals a viable backbone disconnection for split proteins: an approach in identifying a new functional split intein
- PMID: 22937103
- PMCID: PMC3427171
- DOI: 10.1371/journal.pone.0043820
Circular permutation prediction reveals a viable backbone disconnection for split proteins: an approach in identifying a new functional split intein
Abstract
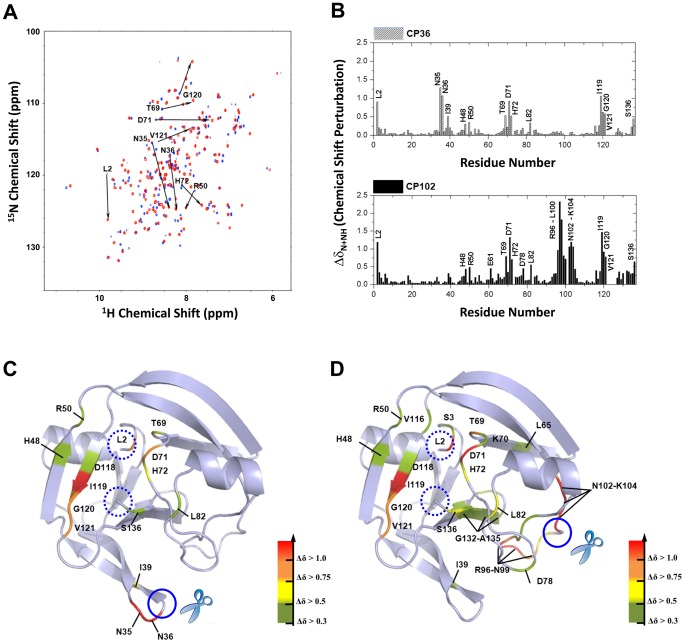
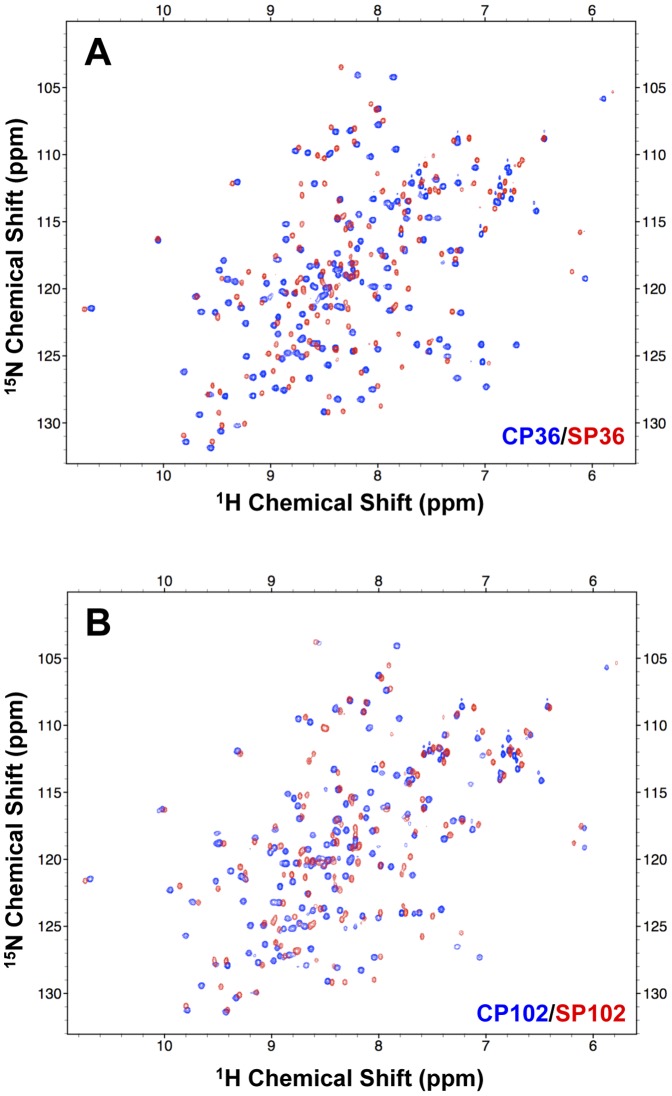
Split-protein systems have emerged as a powerful tool for detecting biomolecular interactions and reporting biological reactions. However, reliable methods for identifying viable split sites are still unavailable. In this study, we demonstrated the feasibility that valid circular permutation (CP) sites in proteins have the potential to act as split sites and that CP prediction can be used to search for internal permissive sites for creating new split proteins. Using a protein ligase, intein, as a model, CP predictor facilitated the creation of circular permutants in which backbone opening imposes the least detrimental effects on intein folding. We screened a series of predicted intein CPs and identified stable and native-fold CPs. When the valid CP sites were introduced as split sites, there was a reduction in folding enthalpy caused by the new backbone opening; however, the coincident loss in entropy was sufficient to be compensated, yielding a favorable free energy for self-association. Since split intein is exploited in protein semi-synthesis, we tested the related protein trans-splicing (PTS) activities of the corresponding split inteins. Notably, a novel functional split intein composed of the N-terminal 36 residues combined with the remaining C-terminal fragment was identified. Its PTS activity was shown to be better than current reported two-piece intein with a short N-terminal segment. Thus, the incorporation of in silico CP prediction facilitated the design of split intein as well as circular permutants.
Conflict of interest statement
Figures






Similar articles
-
Computational Prediction of New Intein Split Sites.Methods Mol Biol. 2017;1495:259-268. doi: 10.1007/978-1-4939-6451-2_17. Methods Mol Biol. 2017. PMID: 27714622
-
A streamlined method for preparing split intein for NMR study.Protein Expr Purif. 2014 Jul;99:106-12. doi: 10.1016/j.pep.2014.04.005. Epub 2014 Apr 19. Protein Expr Purif. 2014. PMID: 24751877
-
Synthetic two-piece and three-piece split inteins for protein trans-splicing.J Biol Chem. 2004 Aug 20;279(34):35281-6. doi: 10.1074/jbc.M405491200. Epub 2004 Jun 11. J Biol Chem. 2004. PMID: 15194682
-
Protein trans-splicing and its use in structural biology: opportunities and limitations.Mol Biosyst. 2010 Nov;6(11):2110-21. doi: 10.1039/c0mb00034e. Epub 2010 Aug 31. Mol Biosyst. 2010. PMID: 20820635 Review.
-
Conditional Split Inteins: Adaptable Tools for Programming Protein Functions.Int J Mol Sci. 2025 Jan 11;26(2):586. doi: 10.3390/ijms26020586. Int J Mol Sci. 2025. PMID: 39859302 Free PMC article. Review.
Cited by
-
Development of an intein-mediated split-Cas9 system for gene therapy.Nucleic Acids Res. 2015 Jul 27;43(13):6450-8. doi: 10.1093/nar/gkv601. Epub 2015 Jun 16. Nucleic Acids Res. 2015. PMID: 26082496 Free PMC article.
-
SeqCP: A sequence-based algorithm for searching circularly permuted proteins.Comput Struct Biotechnol J. 2022 Nov 14;21:185-201. doi: 10.1016/j.csbj.2022.11.024. eCollection 2023. Comput Struct Biotechnol J. 2022. PMID: 36582435 Free PMC article.
-
Nature's recipe for splitting inteins.Protein Eng Des Sel. 2014 Aug;27(8):263-71. doi: 10.1093/protein/gzu028. Protein Eng Des Sel. 2014. PMID: 25096198 Free PMC article. Review.
-
Mitigating genetic instability caused by the excision activity of the phiC31 integrase in Streptomyces.Appl Environ Microbiol. 2025 Jan 31;91(1):e0181224. doi: 10.1128/aem.01812-24. Epub 2024 Dec 20. Appl Environ Microbiol. 2025. PMID: 39704534 Free PMC article.
-
SPLICEFINDER - a fast and easy screening method for active protein trans-splicing positions.PLoS One. 2013 Sep 2;8(9):e72925. doi: 10.1371/journal.pone.0072925. eCollection 2013. PLoS One. 2013. PMID: 24023792 Free PMC article.
References
-
- Yu Y, Lutz S (2011) Circular permutation: a different way to engineer enzyme structure and function. Trends in biotechnology 29: 18–25. - PubMed
-
- Heinemann U, Hahn M (1995) Circular permutation of polypeptide chains: implications for protein folding and stability. Progress in biophysics and molecular biology 64: 121–143. - PubMed
-
- Lindqvist Y, Schneider G (1997) Circular permutations of natural protein sequences: structural evidence. Current opinion in structural biology 7: 422–427. - PubMed
-
- Whitehead TA, Bergeron LM, Clark DS (2009) Tying up the loose ends: circular permutation decreases the proteolytic susceptibility of recombinant proteins. Protein engineering, design & selection : PEDS 22: 607–613. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
