

Neuroprotection after stroke by targeting NOX4 as a source of oxidative stress
- PMID: 22937798
- PMCID: PMC3603500
- DOI: 10.1089/ars.2012.4797
Neuroprotection after stroke by targeting NOX4 as a source of oxidative stress
Abstract
Significance: Stroke, a leading cause of death and disability, poses a substantial burden for patients, relatives, and our healthcare systems. Only one drug is approved for treating stroke, and more than 30 contraindications exclude its use in 90% of all patients. Thus, new treatments are urgently needed. In this review, we discuss oxidative stress as a pathomechanism of poststroke neurodegeneration and the inhibition of its source, type 4 nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX4), as a conceptual breakthrough in stroke therapy.
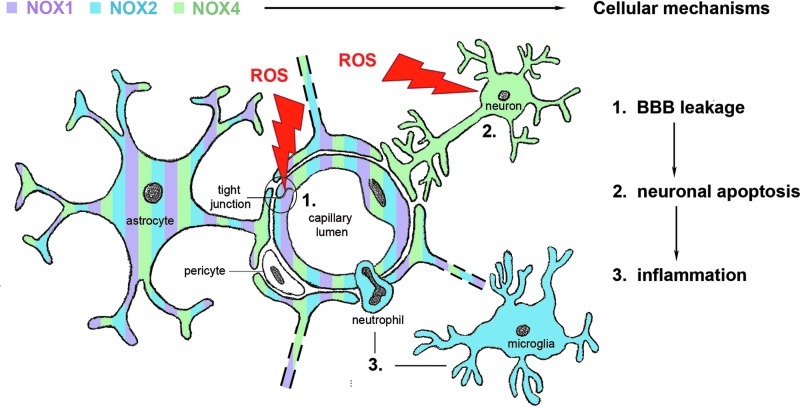
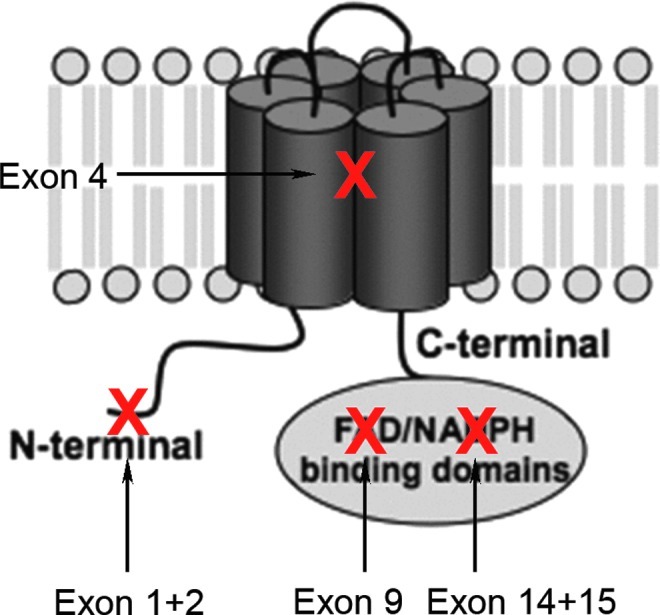
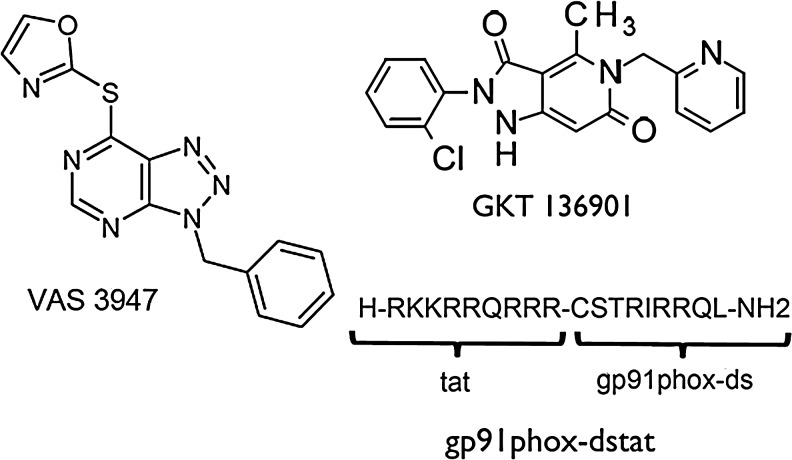
Recent advances: Among potential sources of reactive oxygen species (ROS), the NOXes stand out as the only enzyme family that is solely dedicated to forming ROS. In rodents, three cerebrovascular NOXes exist: the superoxide-forming NOX1 and 2 and the hydrogen peroxide-forming NOX4. Studies using NOX1 knockout mice gave conflicting results, which overall do not point to a role for this isoform. Several reports find NOX2 to be relevant in stroke, albeit to variable and moderate degrees. In our hands, NOX4 is, by far, the major source of oxidative stress and neurodegeneration on ischemic stroke.
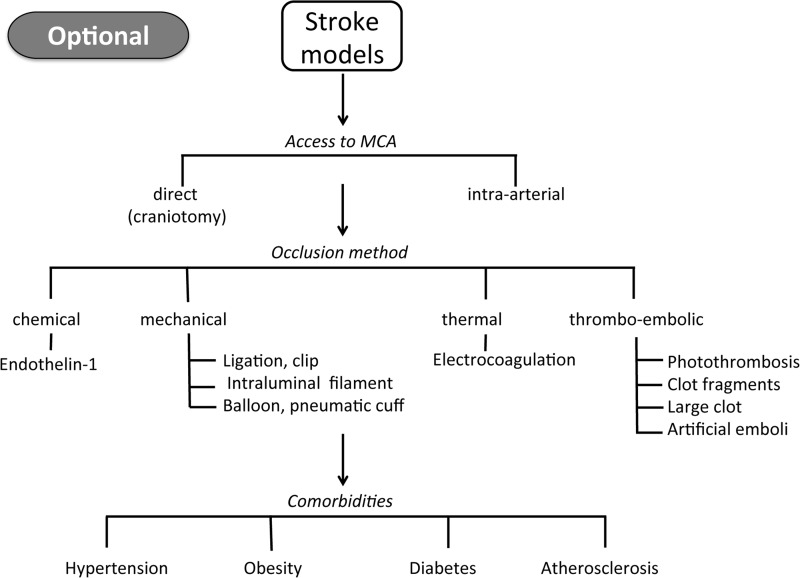
Critical issues: We critically discuss the tools that have been used to validate the roles of NOX in stroke. We also highlight the relevance of different animal models and the need for advanced quality control in preclinical stroke research.
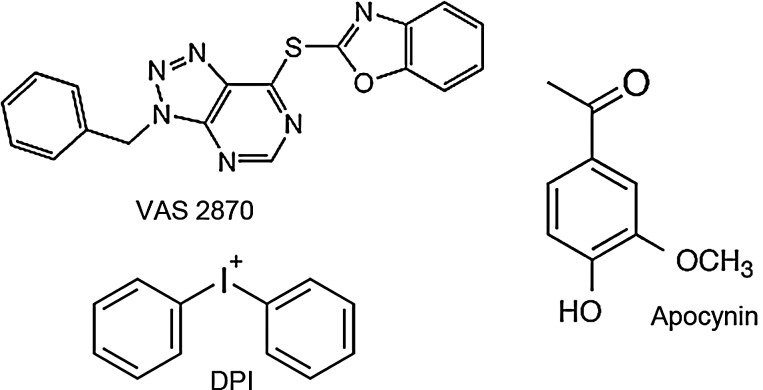
Future directions: The development of isoform-specific NOX inhibitors presents a precious tool for further clarifying the role and drugability of NOX homologues. This could pave the avenue for the first clinically effective neuroprotectant applied poststroke, and even beyond this, stroke could provide a proof of principle for antioxidative stress therapy.
Figures

References
-
- Ago T. Kitazono T. Kuroda J. Kumai Y. Kamouchi M. Ooboshi H. Wakisaka M. Kawahara T. Rokutan K. Ibayashi S. Iida M. NAD(P)H oxidases in rat basilar arterial endothelial cells. Stroke. 2005;36:1040–1046. - PubMed
-
- Aldieri E. Riganti C. Polimeni M. Gazzano E. Lussiana C. Campia I. Ghigo D. Classical inhibitors of NOX NAD(P)H oxidases are not specific. Curr Drug Metab. 2008;9:686–696. - PubMed
-
- Anilkumar N. Weber R. Zhang M. Brewer A. Shah AM. Nox4 and nox2 NADPH oxidases mediate distinct cellular redox signaling responses to agonist stimulation. Arterioscler Thromb Vasc Biol. 2008;28:1347–1354. - PubMed
-
- Arimura K. Ago T. Kuroda J. Ishitsuka K. Nishimura A. Sugimori H. Kamouchi M. Sasaki T. Kitazono T. Role of NADPH oxidase 4 in Brain Endothelial cells after Ischemic Stroke. Stroke. 2012;43:A2514.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
