

Endocytosis of the seven-transmembrane RGS1 protein activates G-protein-coupled signalling in Arabidopsis
- PMID: 22940907
- PMCID: PMC3463750
- DOI: 10.1038/ncb2568
Endocytosis of the seven-transmembrane RGS1 protein activates G-protein-coupled signalling in Arabidopsis
Abstract
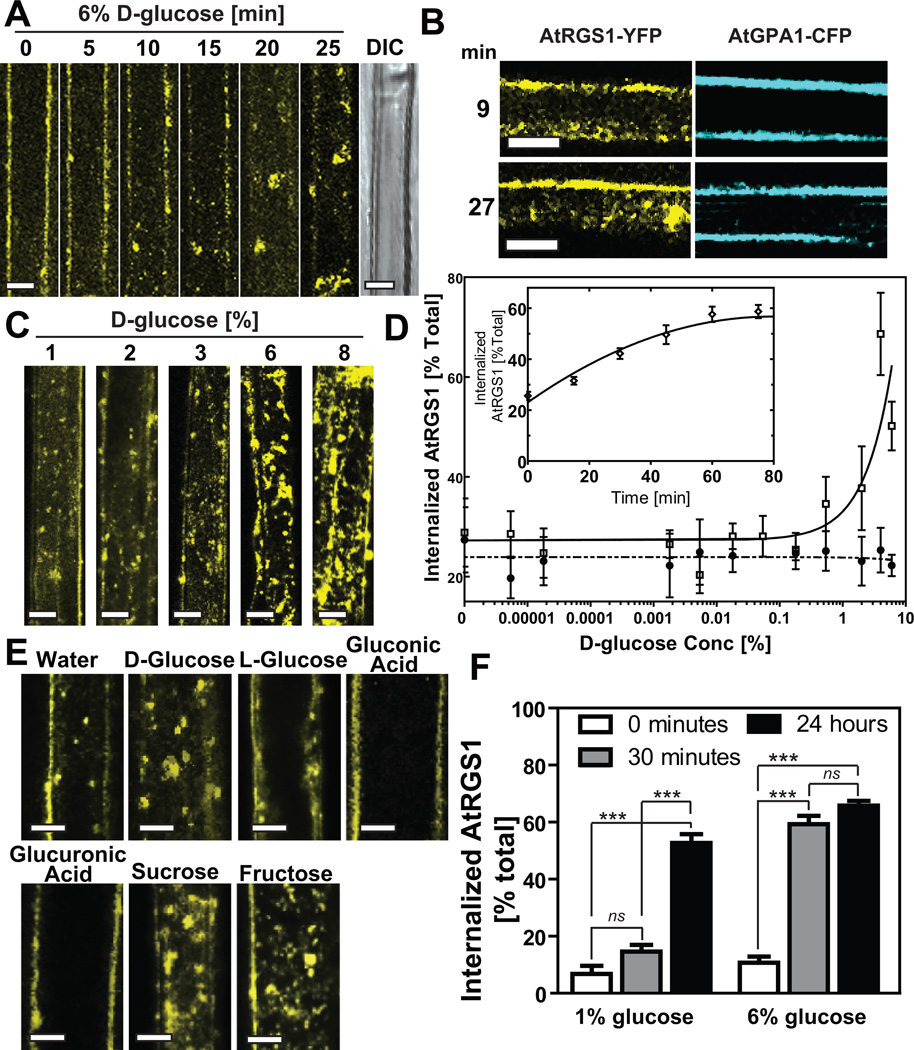
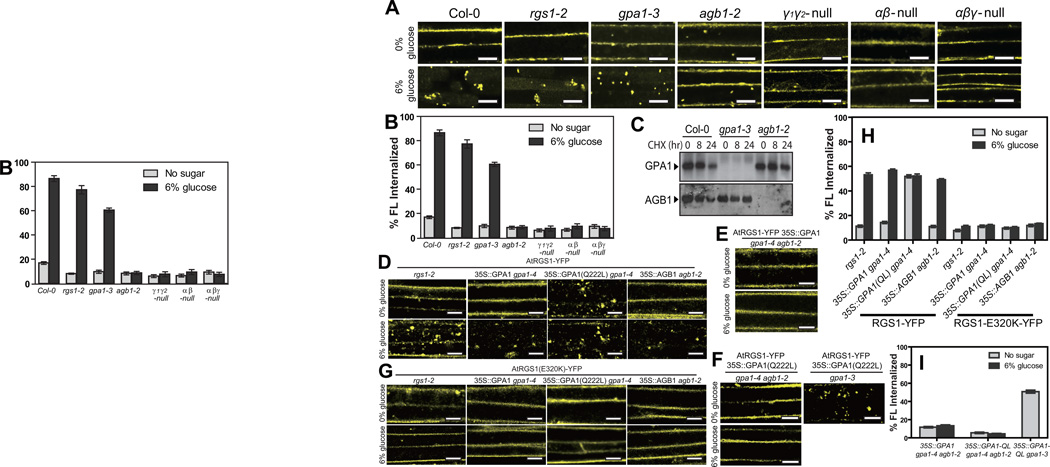
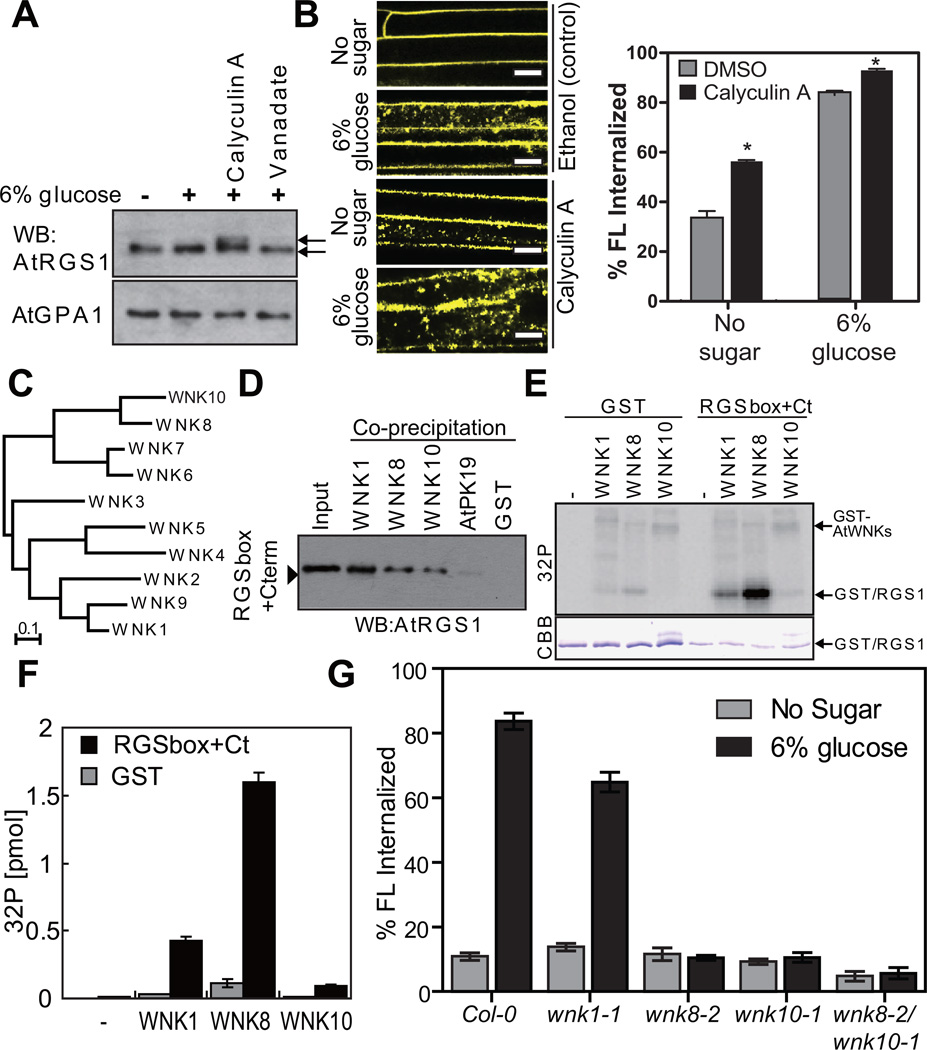
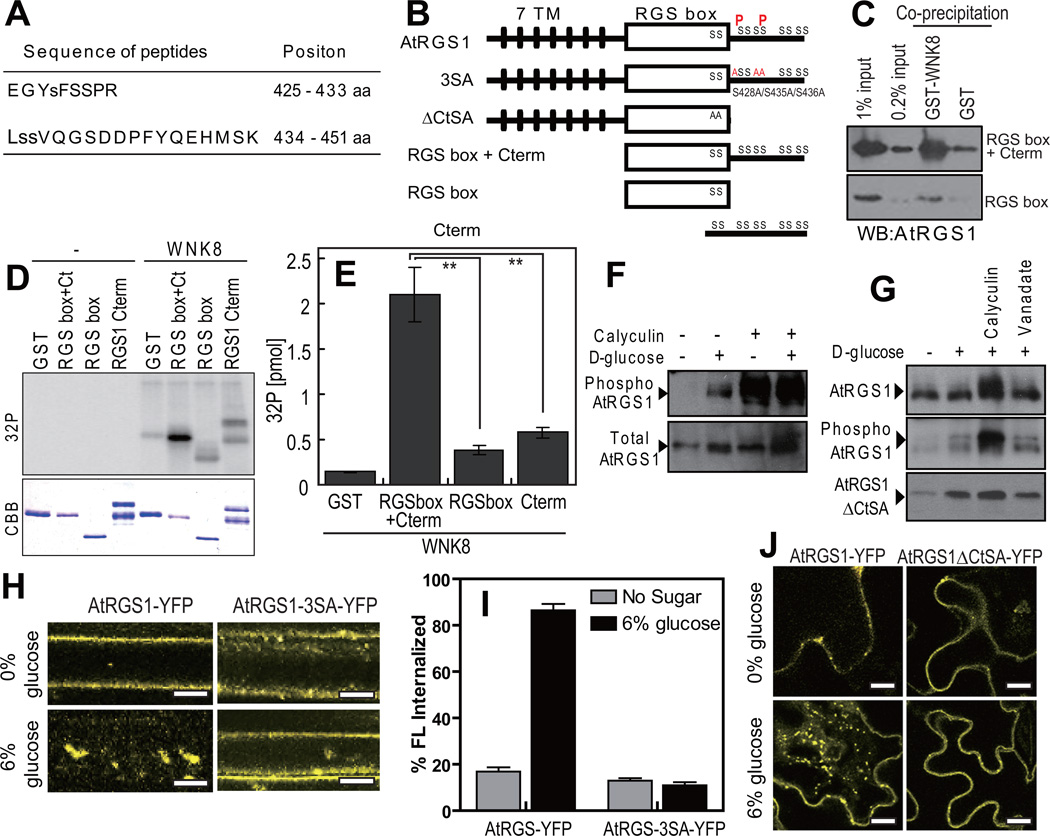
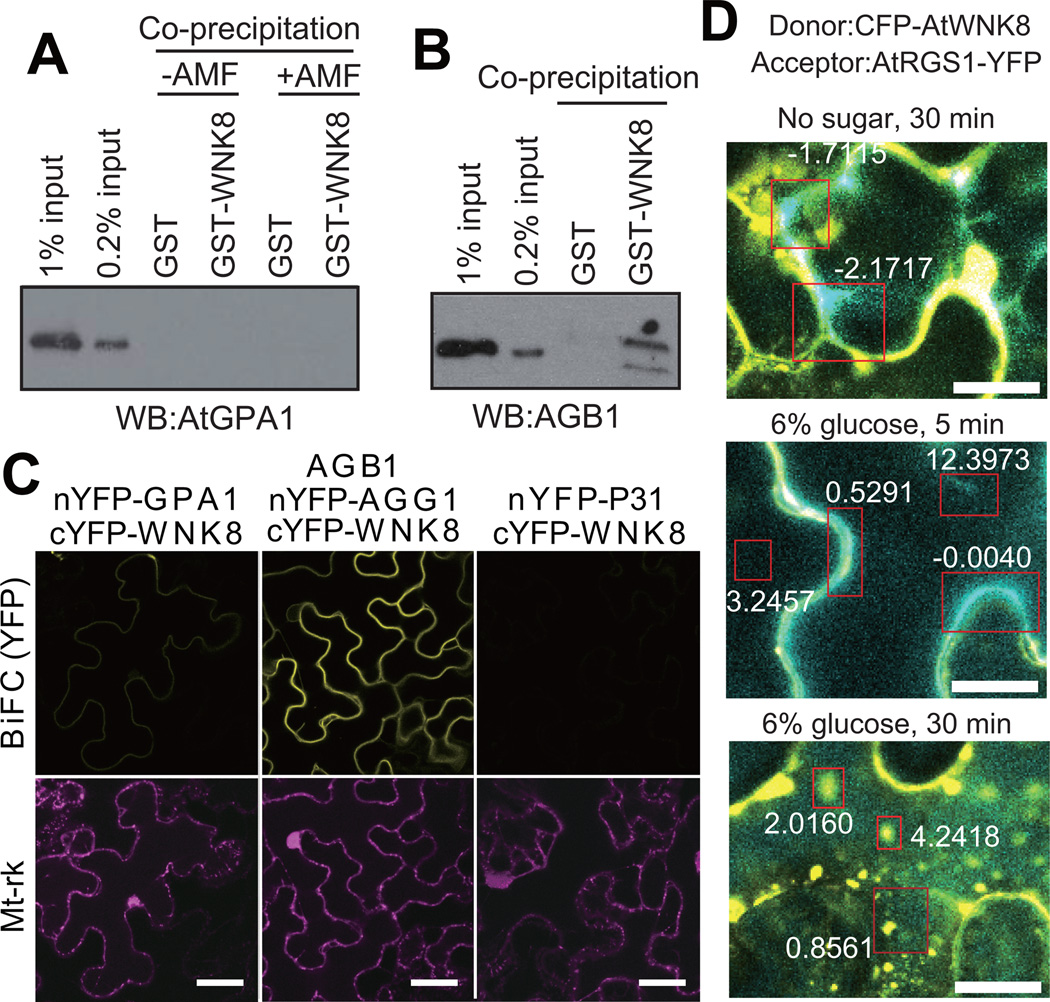
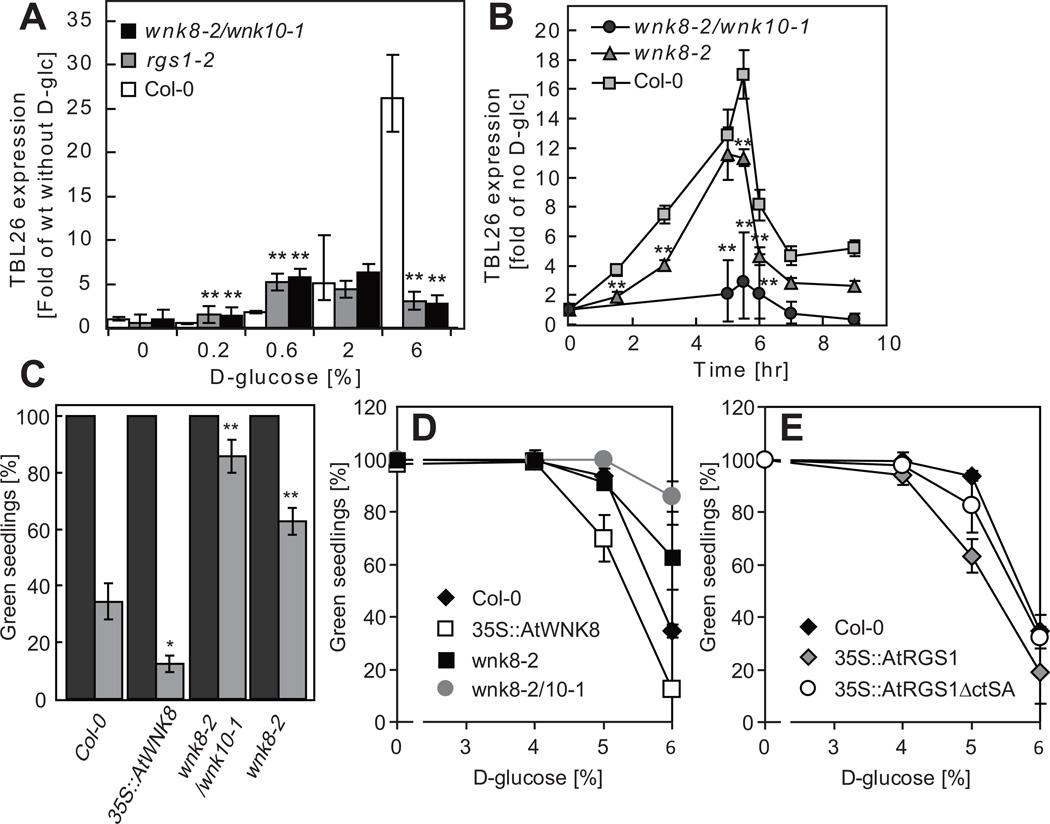
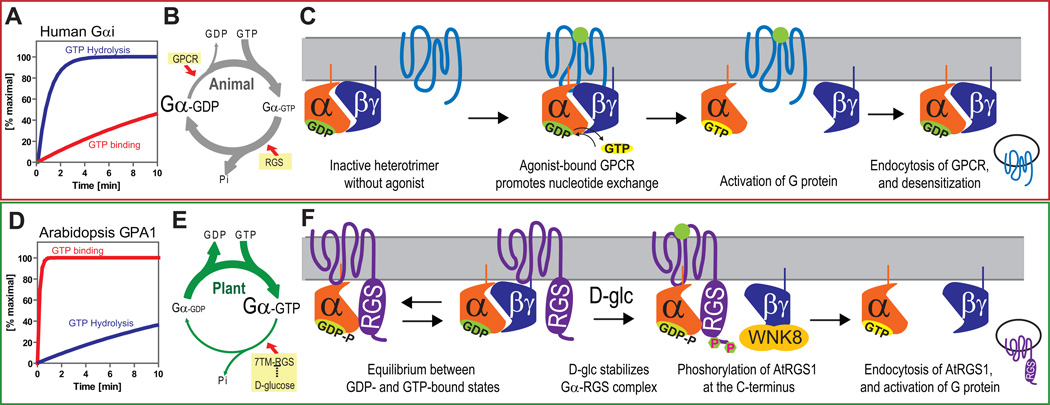
Signal transduction typically begins by ligand-dependent activation of a concomitant partner that is otherwise in its resting state. However, in cases where signal activation is constitutive by default, the mechanism of regulation is unknown. The Arabidopsis thaliana heterotrimeric Gα protein self-activates without accessory proteins, and is kept in its resting state by the negative regulator, AtRGS1 (regulator of G-protein signalling 1), which is the prototype of a seven-transmembrane receptor fused with an RGS domain. Endocytosis of AtRGS1 by ligand-dependent endocytosis physically uncouples the GTPase-accelerating activity of AtRGS1 from the Gα protein, permitting sustained activation. Phosphorylation of AtRGS1 by AtWNK8 kinase causes AtRGS1 endocytosis, required for both G-protein-mediated sugar signalling and cell proliferation. In animals, receptor endocytosis results in signal desensitization, whereas in plants, endocytosis results in signal activation. These findings reveal how different organisms rearrange a regulatory system to result in opposite outcomes using similar phosphorylation-dependent endocytosis mechanisms.
Figures
References
-
- Kohout TA, Lefkowitz RJ. Regulation of G protein-coupled receptor kinases and arrestins during receptor desensitization. Mol Pharmacol. 2003;63:9–18. - PubMed
-
- Hanyaloglu AC, von Zastrow M. Regulation of GPCRs by endocytic membrane trafficking and its potential implications. Annu Rev Pharmacol Toxicol. 2008;48:537–568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
