

EG-VEGF controls placental growth and survival in normal and pathological pregnancies: case of fetal growth restriction (FGR)
- PMID: 22941044
- PMCID: PMC11113665
- DOI: 10.1007/s00018-012-1141-z
EG-VEGF controls placental growth and survival in normal and pathological pregnancies: case of fetal growth restriction (FGR)
Abstract
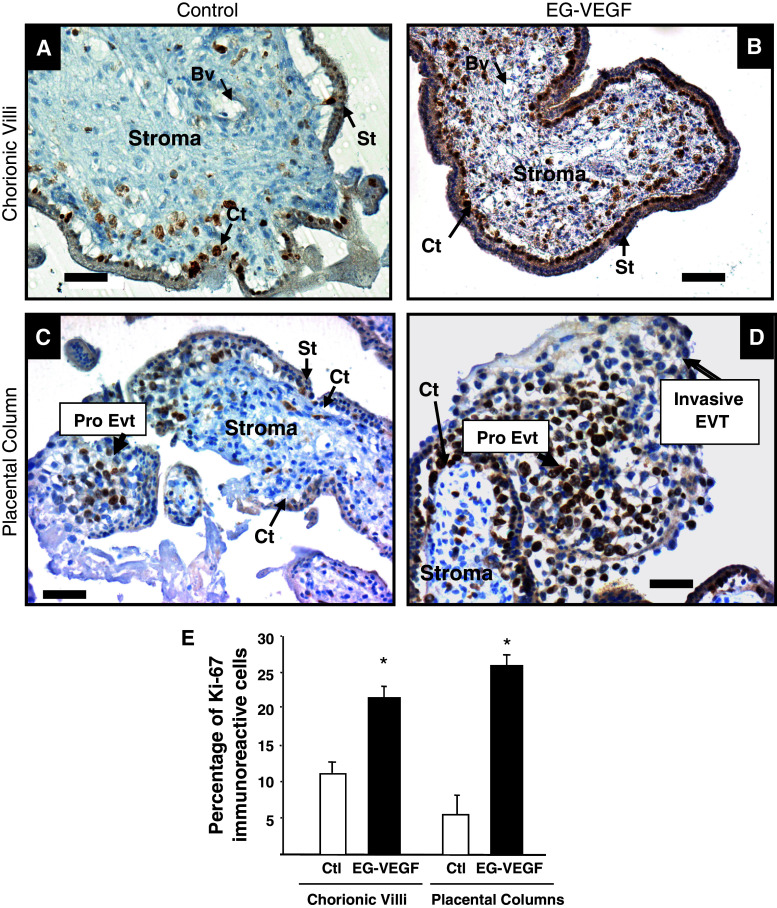
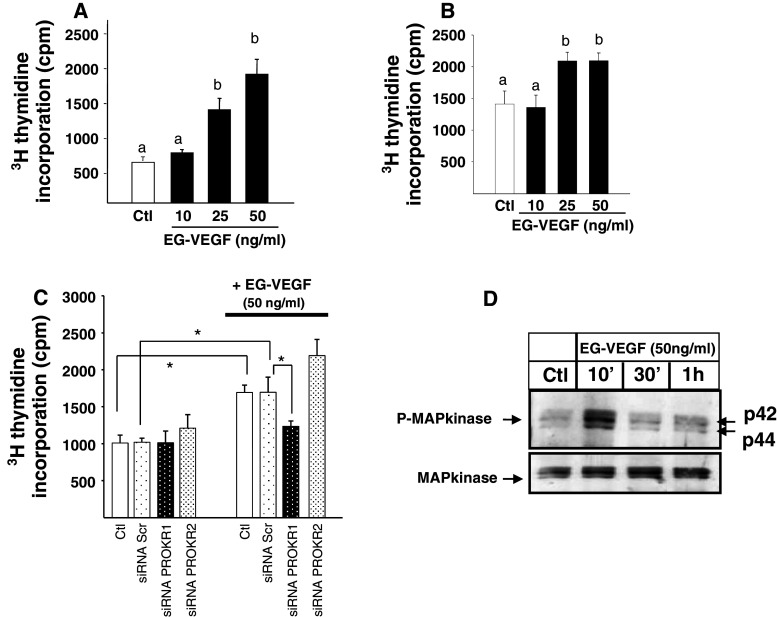
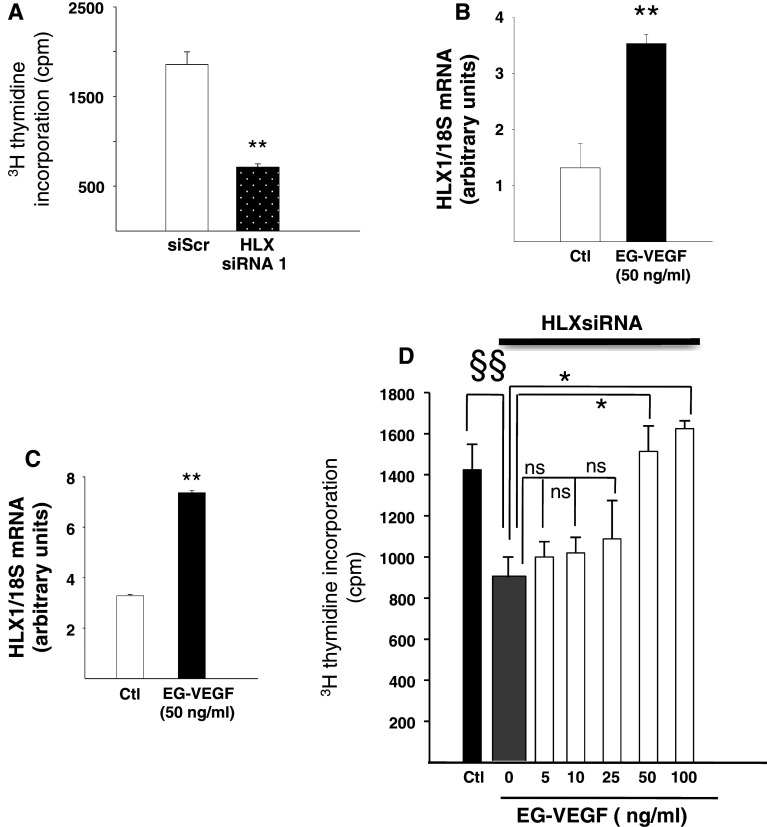
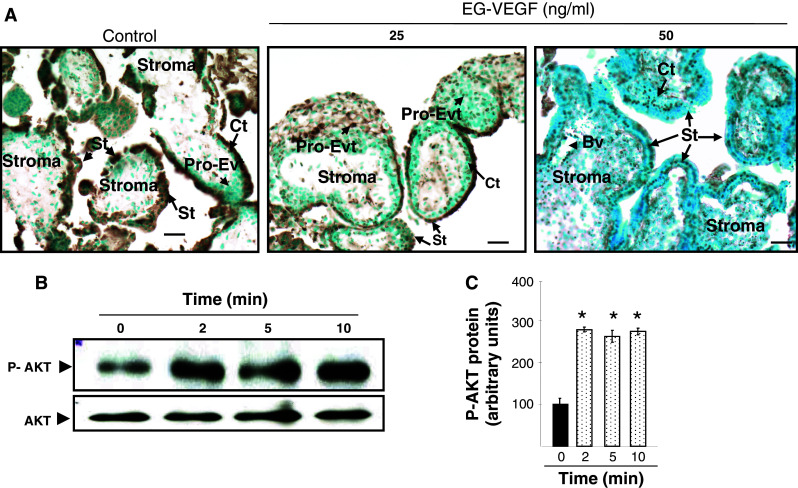
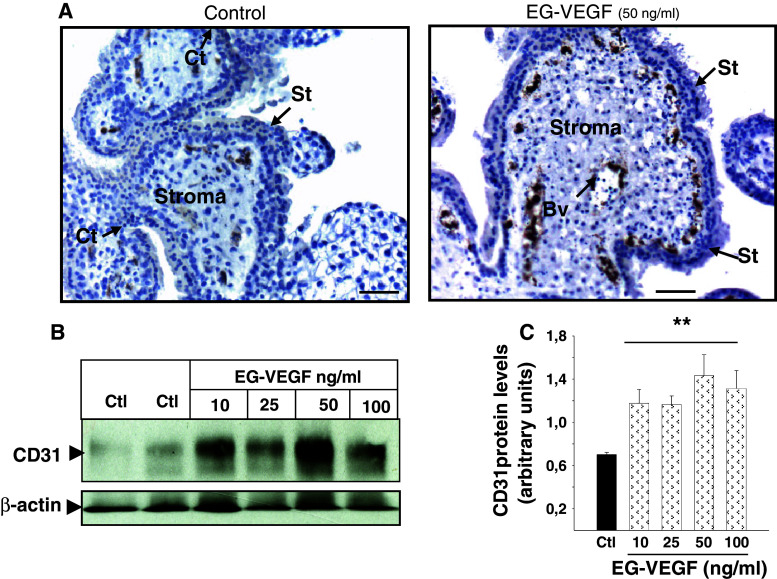
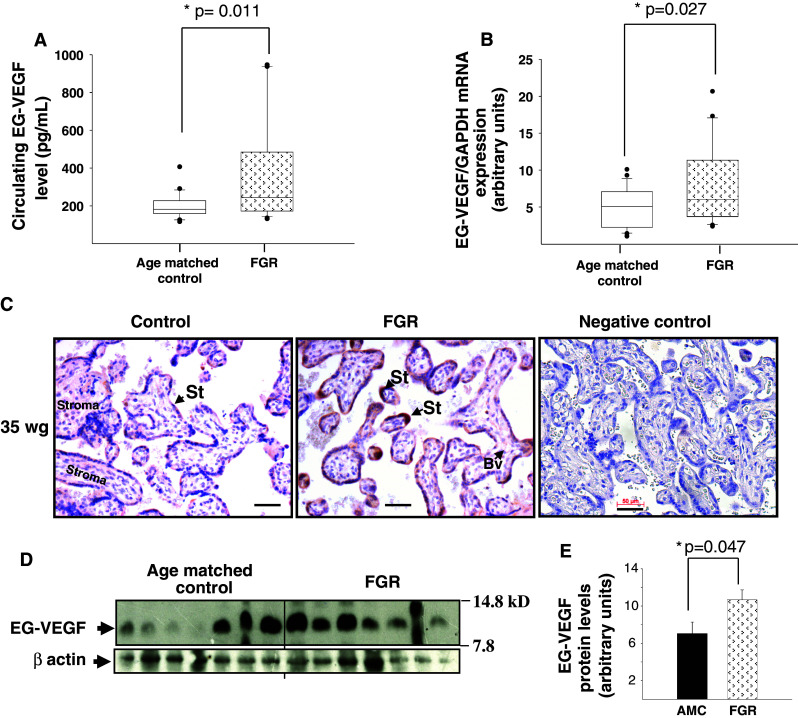
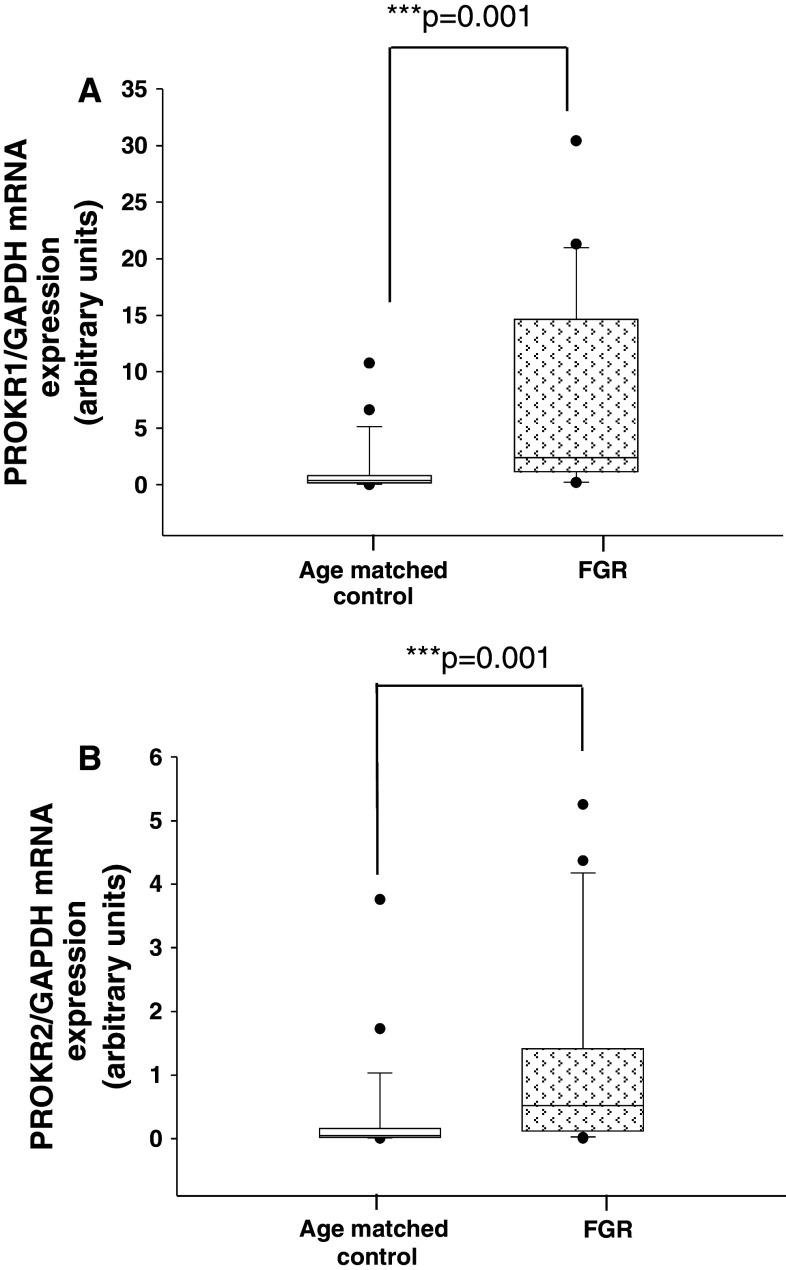
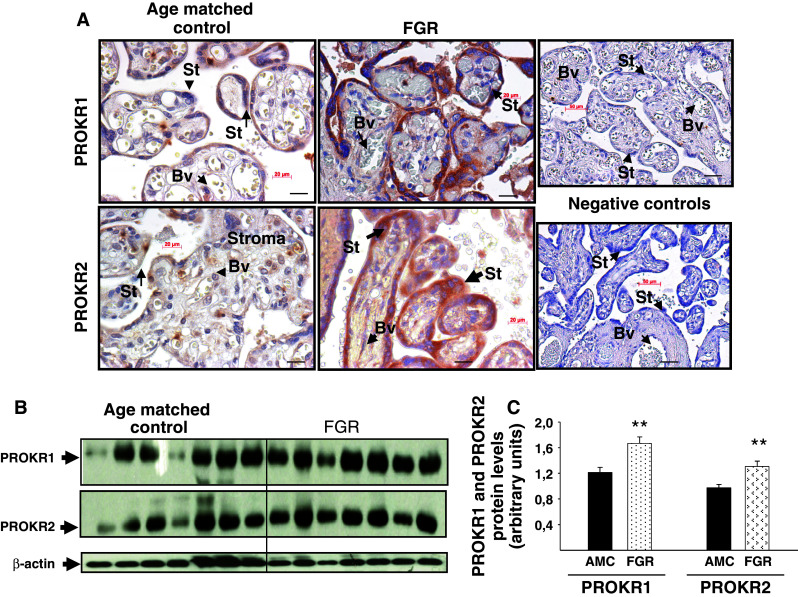
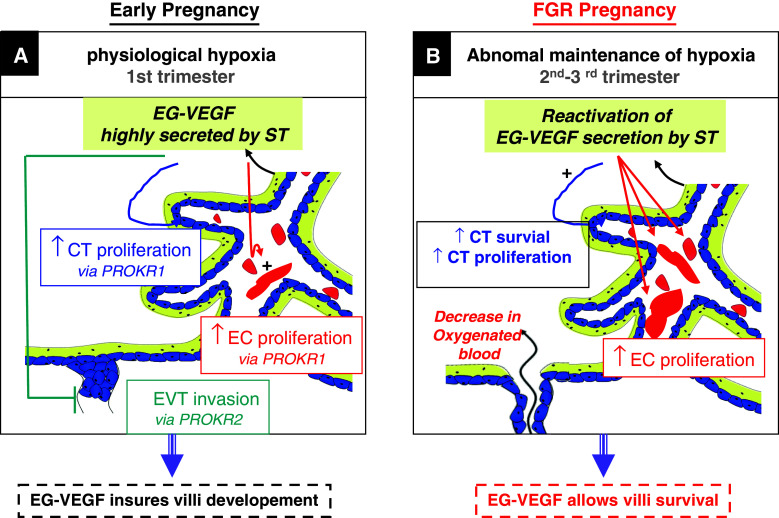
Identifiable causes of fetal growth restriction (FGR) account for 30 % of cases, but the remainders are idiopathic and are frequently associated with placental dysfunction. We have shown that the angiogenic factor endocrine gland-derived VEGF (EG-VEGF) and its receptors, prokineticin receptor 1 (PROKR1) and 2, (1) are abundantly expressed in human placenta, (2) are up-regulated by hypoxia, (3) control trophoblast invasion, and that EG-VEGF circulating levels are the highest during the first trimester of pregnancy, the period of important placental growth. These findings suggest that EG-VEGF/PROKR1 and 2 might be involved in normal and FGR placental development. To test this hypothesis, we used placental explants, primary trophoblast cultures, and placental and serum samples collected from FGR and age-matched control women. Our results show that (1) EG-VEGF increases trophoblast proliferation ([(3)H]-thymidine incorporation and Ki67-staining) via the homeobox-gene, HLX (2) the proliferative effect involves PROKR1 but not PROKR2, (3) EG-VEGF does not affect syncytium formation (measurement of syncytin 1 and 2 and β hCG production) (4) EG-VEGF increases the vascularization of the placental villi and insures their survival, (5) EG-VEGF, PROKR1, and PROKR2 mRNA and protein levels are significantly elevated in FGR placentas, and (6) EG-VEGF circulating levels are significantly higher in FGR patients. Altogether, our results identify EG-VEGF as a new placental growth factor acting during the first trimester of pregnancy, established its mechanism of action, and provide evidence for its deregulation in FGR. We propose that EG-VEGF/PROKR1 and 2 increases occur in FGR as a compensatory mechanism to insure proper pregnancy progress.
Figures
References
-
- Cross JC, Baczyk D, Dobric N, Hemberger M, Hughes M, Simmons DG, Yamamoto H, Kingdom JC. Genes, development and evolution of the placenta. Placenta. 2003;24(2–3):123–130. - PubMed
-
- Henriksen T. Foetal nutrition, foetal growth restriction and health later in life. Acta Paediatr Suppl. 1999;88(429):4–8. - PubMed
-
- Barker DJ. Fetal growth and adult disease. Br J Obstet Gynaecol. 1992;99(4):275–276. - PubMed
-
- Bernstein IM, Horbar JD, Badger GJ, Ohlsson A, Golan A. Morbidity and mortality among very-low-birth-weight neonates with intrauterine growth restriction. The Vermont Oxford Network. Am J Obstet Gynecol. 2000;182(1 Pt 1):198–206. - PubMed
-
- Baschat AA. Fetal responses to placental insufficiency: an update. BJOG. 2004;111(10):1031–1041. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
