

The physical state of lipid substrates provides transacylation specificity for tafazzin
- PMID: 22941046
- PMCID: PMC3699345
- DOI: 10.1038/nchembio.1064
The physical state of lipid substrates provides transacylation specificity for tafazzin
Abstract
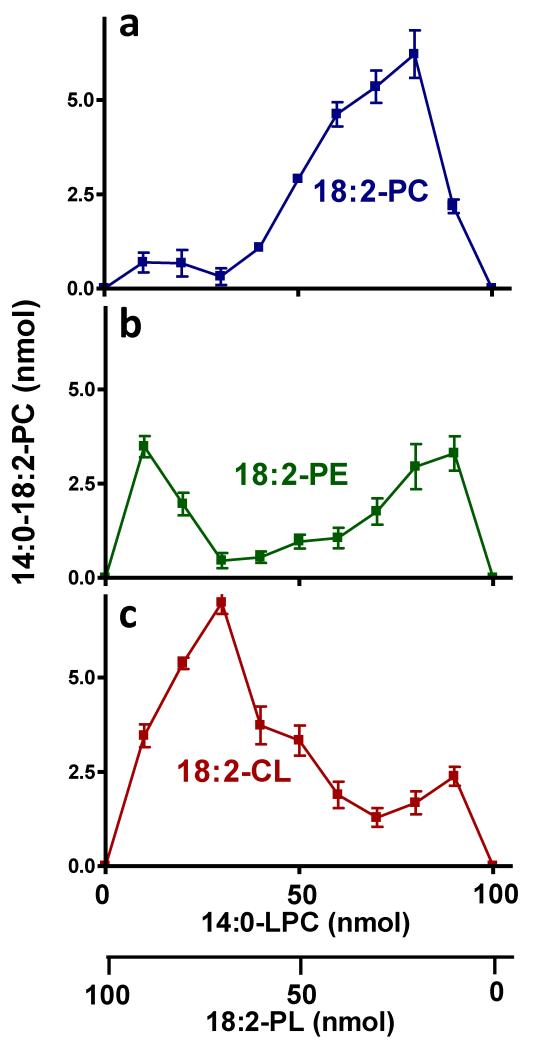
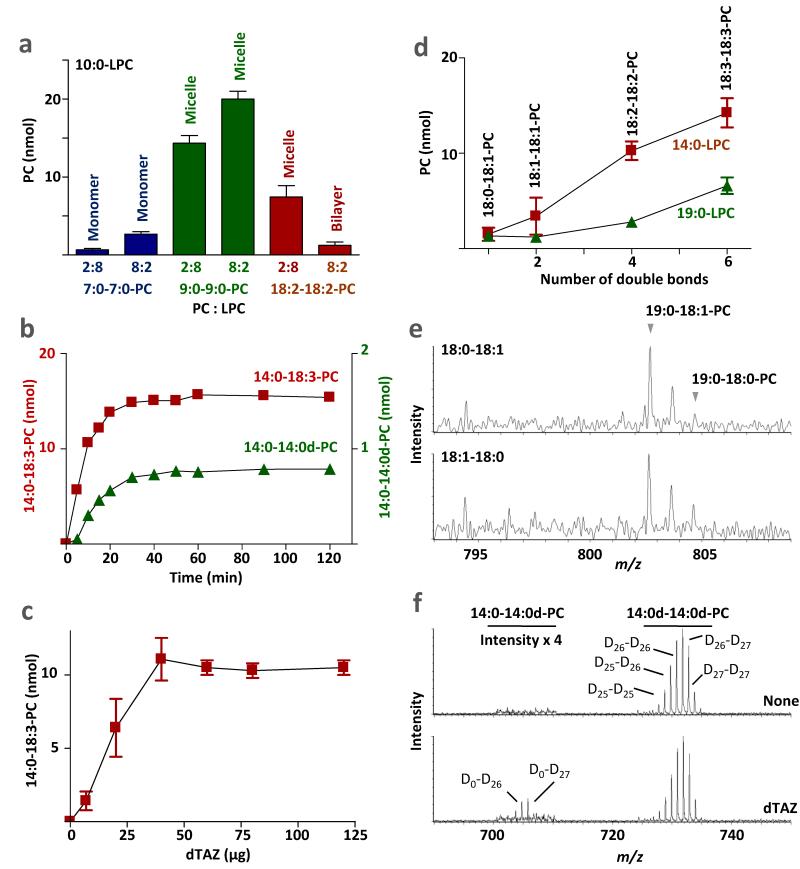
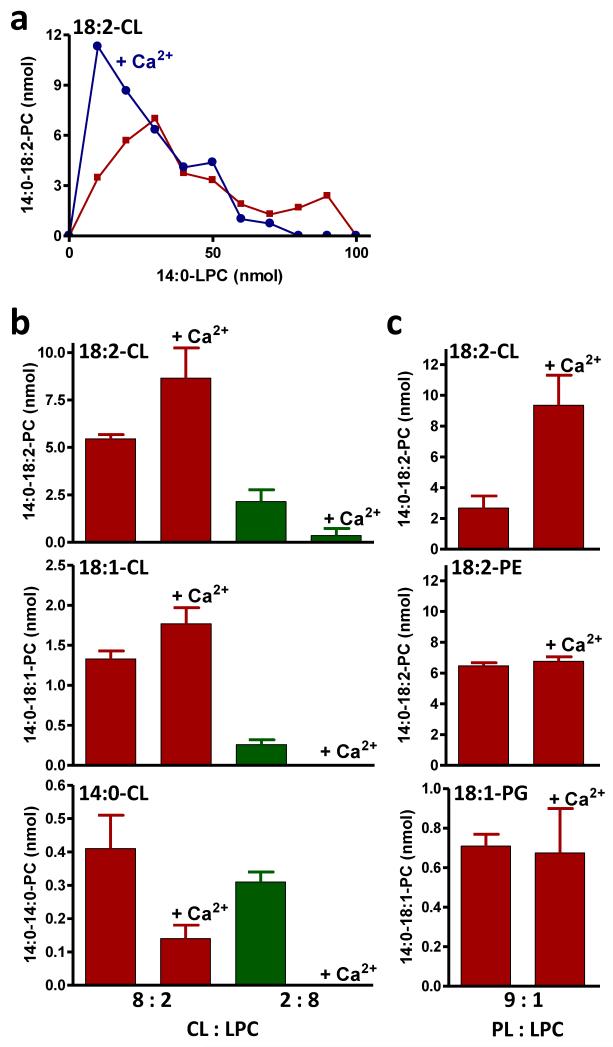
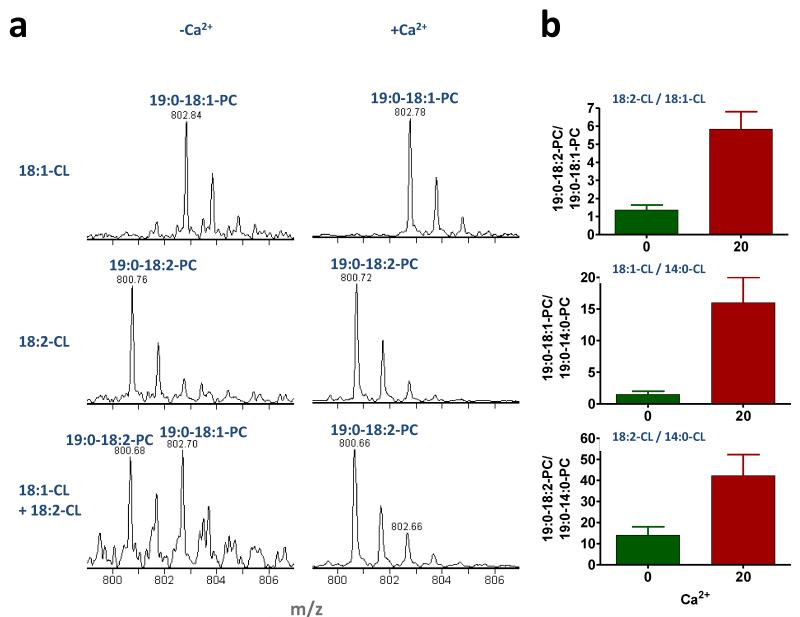
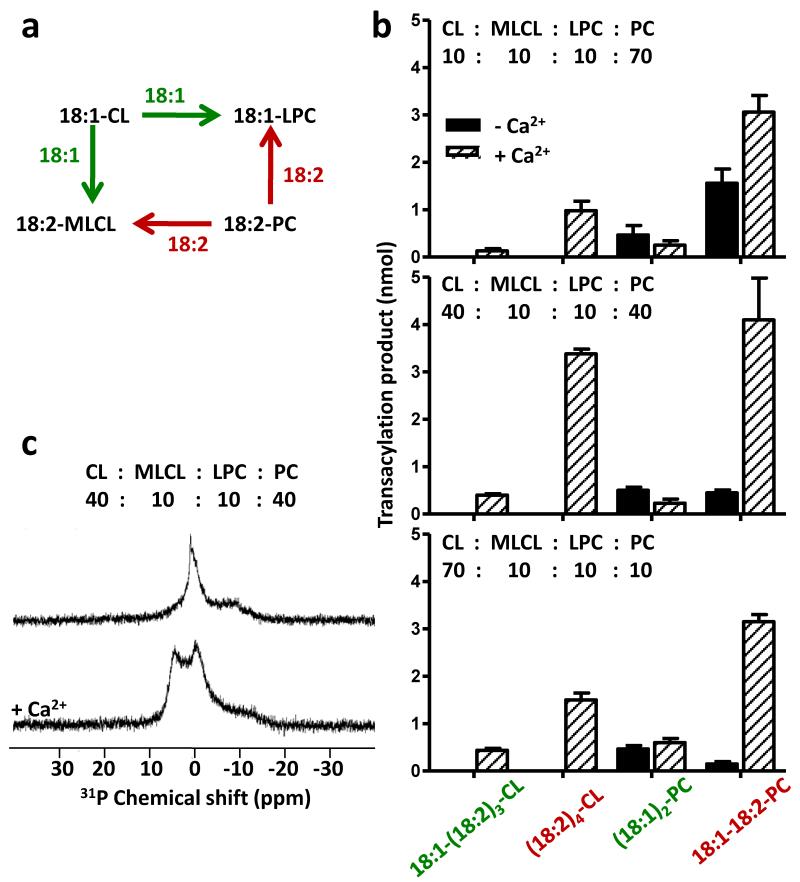
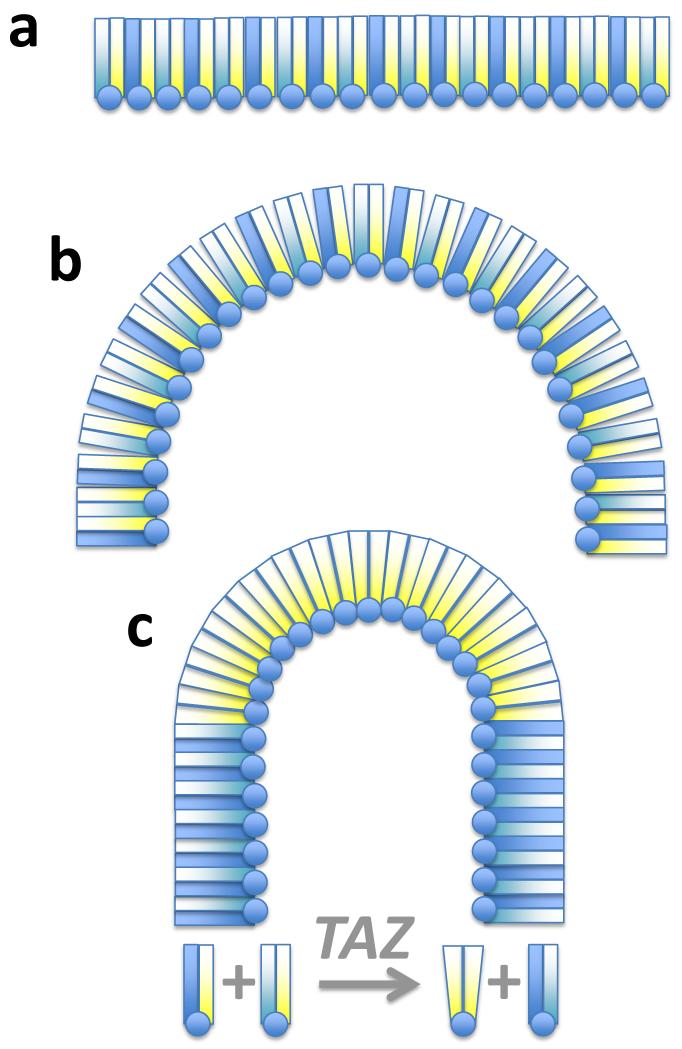
Cardiolipin is a mitochondrial phospholipid with a characteristic acyl chain composition that depends on the function of tafazzin, a phospholipid-lysophospholipid transacylase, although the enzyme itself lacks acyl specificity. We incubated isolated tafazzin with various mixtures of phospholipids and lysophospholipids, characterized the lipid phase by (31)P-NMR and measured newly formed molecular species by MS. Substantial transacylation was observed only in nonbilayer lipid aggregates, and the substrate specificity was highly sensitive to the lipid phase. In particular, tetralinoleoyl-cardiolipin, a prototype molecular species, formed only under conditions that favor the inverted hexagonal phase. In isolated mitochondria, <1% of lipids participated in transacylations, suggesting that the action of tafazzin was limited to privileged lipid domains. We propose that tafazzin reacts with non-bilayer-type lipid domains that occur in curved or hemifused membrane zones and that acyl specificity is driven by the packing properties of these domains.
Figures
Comment in
-
Tafazzin senses curvature.Nat Chem Biol. 2012 Oct;8(10):811-2. doi: 10.1038/nchembio.1068. Nat Chem Biol. 2012. PMID: 22987008 Free PMC article.
References
-
- Ma L, Vaz FM, Gu Z, Wanders RJA, Greenberg ML. The human TAZ gene complements mitochondrial dysfunction in the yeast taz1Δ mutant. Implications for Barth syndrome. J. Biol. Chem. 2004;279:44394–44399. - PubMed
-
- Xu Y, Malhotra A, Ren M, Schlame M. The enzymatic function of tafazzin. J. Biol. Chem. 2006;281:39217–39224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
