

Calcium entry induces mitochondrial oxidant stress in vagal neurons at risk in Parkinson's disease
- PMID: 22941107
- PMCID: PMC3461271
- DOI: 10.1038/nn.3209
Calcium entry induces mitochondrial oxidant stress in vagal neurons at risk in Parkinson's disease
Abstract
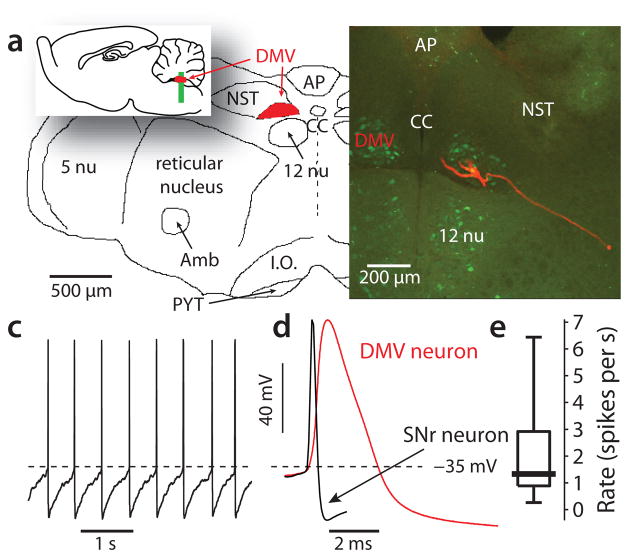
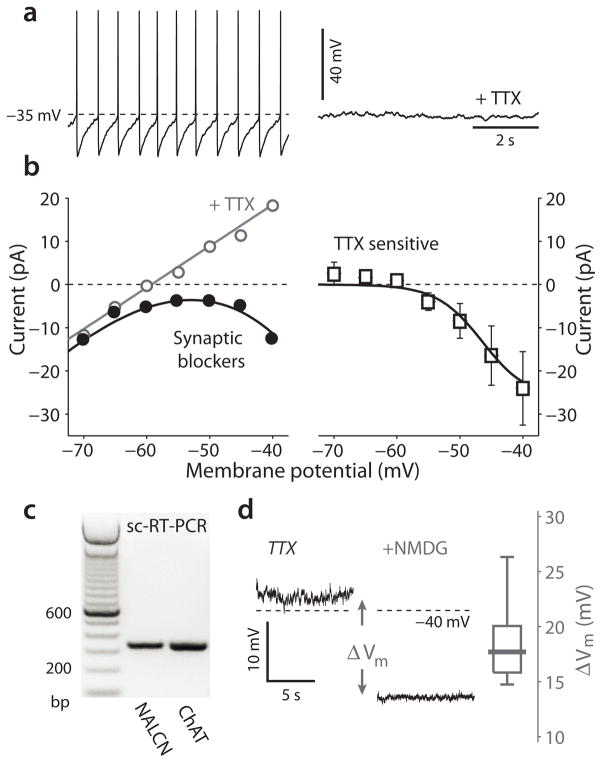
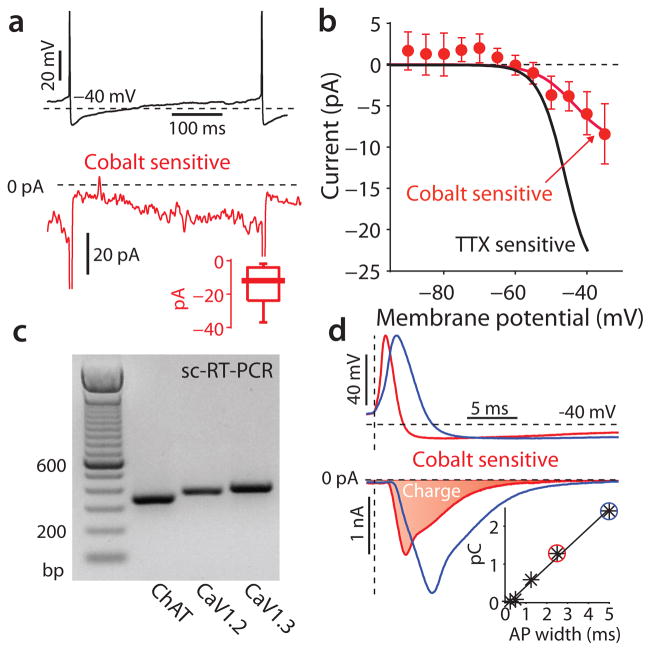
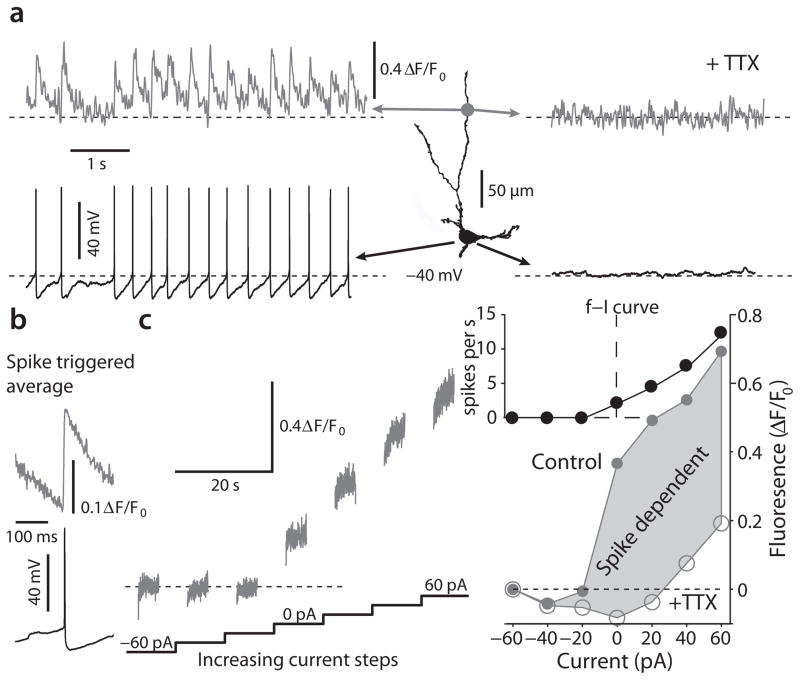
Mitochondrial oxidant stress is widely viewed as being critical to pathogenesis in Parkinson's disease. But the origins of this stress are poorly defined. One possibility is that it arises from the metabolic demands associated with regenerative activity. To test this hypothesis, we characterized neurons in the dorsal motor nucleus of the vagus (DMV), a population of cholinergic neurons that show signs of pathology in the early stages of Parkinson's disease, in mouse brain slices. DMV neurons were slow, autonomous pacemakers with broad spikes, leading to calcium entry that was weakly buffered. Using a transgenic mouse expressing a redox-sensitive optical probe targeted to the mitochondrial matrix, we found that calcium entry during pacemaking created a basal mitochondrial oxidant stress. Knocking out DJ-1 (also known as PARK7), a gene associated with early-onset Parkinson's disease, exacerbated this stress. These results point to a common mechanism underlying mitochondrial oxidant stress in Parkinson's disease and a therapeutic strategy to ameliorate it.
Conflict of interest statement
Competing financial interests
The authors declare no competing financial interests.
Figures
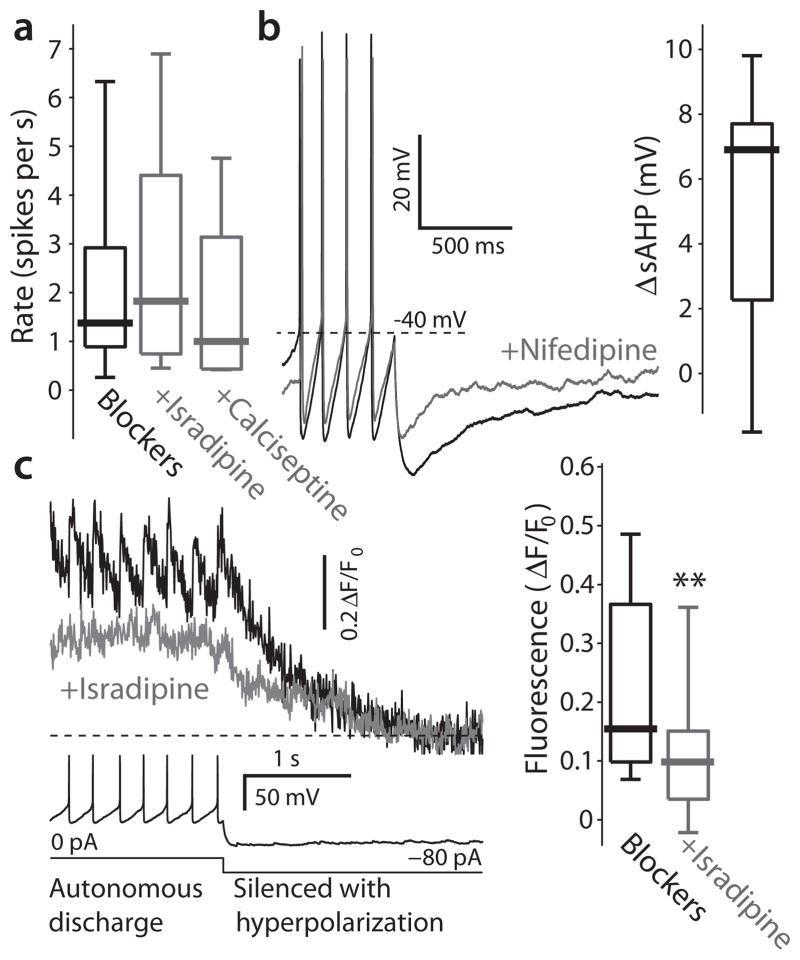
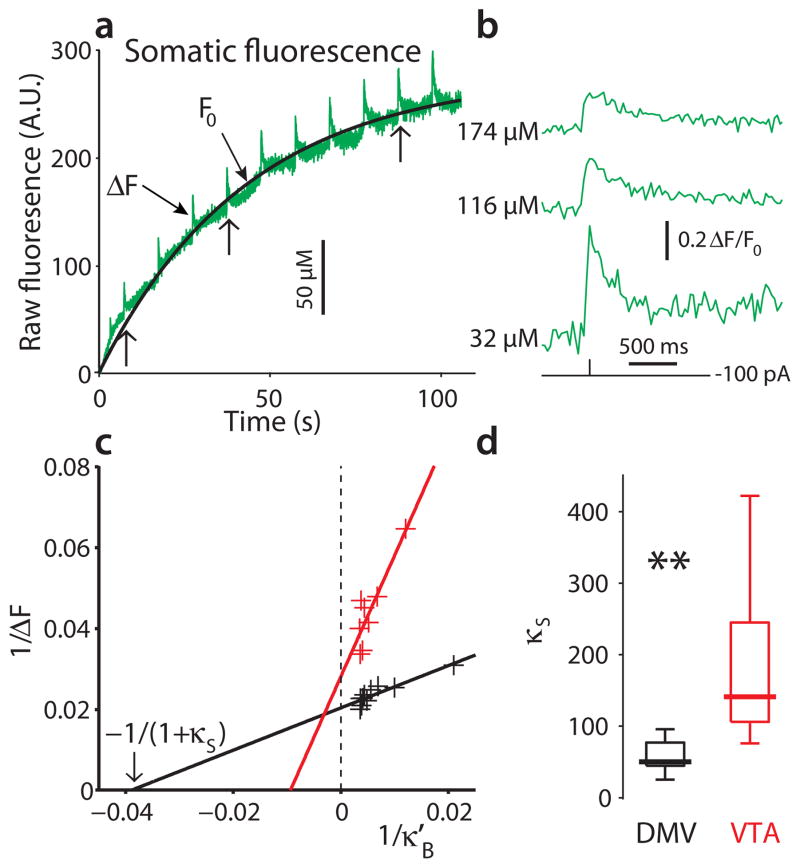
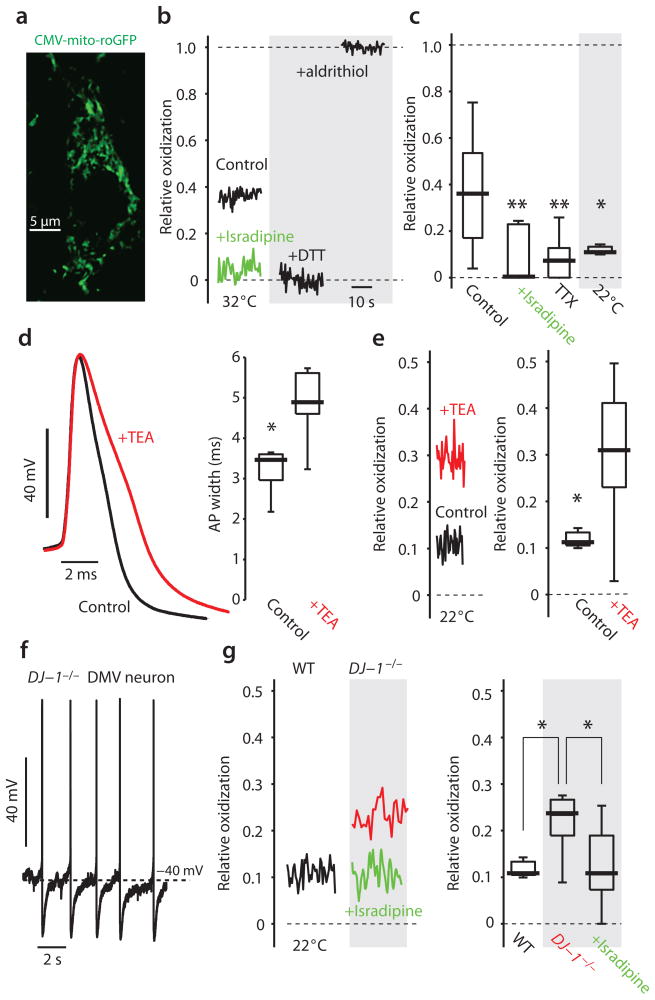
Comment in
-
Calcium as main trigger of mitochondrial oxidant stress in Parkinson's disease.Mov Disord. 2013 Mar;28(3):288. doi: 10.1002/mds.25331. Mov Disord. 2013. PMID: 23526427 No abstract available.
Similar articles
-
Oxidant stress evoked by pacemaking in dopaminergic neurons is attenuated by DJ-1.Nature. 2010 Dec 2;468(7324):696-700. doi: 10.1038/nature09536. Epub 2010 Nov 10. Nature. 2010. PMID: 21068725 Free PMC article.
-
Mutant α-Synuclein Overexpression Induces Stressless Pacemaking in Vagal Motoneurons at Risk in Parkinson's Disease.J Neurosci. 2017 Jan 4;37(1):47-57. doi: 10.1523/JNEUROSCI.1079-16.2016. J Neurosci. 2017. PMID: 28053029 Free PMC article.
-
DJ-1 mediates paraquat-induced dopaminergic neuronal cell death.Toxicol Lett. 2011 Apr 25;202(2):85-92. doi: 10.1016/j.toxlet.2011.01.018. Epub 2011 Feb 21. Toxicol Lett. 2011. PMID: 21300143
-
[Mitochondrial dysfunction and oxidative damages in the molecular pathology of Parkinson's disease].Mol Biol (Mosk). 2008 Sep-Oct;42(5):809-19. Mol Biol (Mosk). 2008. PMID: 18988530 Review. Russian.
-
Pink1, Parkin, DJ-1 and mitochondrial dysfunction in Parkinson's disease.Curr Opin Neurobiol. 2007 Jun;17(3):331-7. doi: 10.1016/j.conb.2007.04.010. Epub 2007 May 11. Curr Opin Neurobiol. 2007. PMID: 17499497 Review.
Cited by
-
Metformin Protects From Rotenone-Induced Nigrostriatal Neuronal Death in Adult Mice by Activating AMPK-FOXO3 Signaling and Mitigation of Angiogenesis.Front Mol Neurosci. 2020 Jun 18;13:84. doi: 10.3389/fnmol.2020.00084. eCollection 2020. Front Mol Neurosci. 2020. PMID: 32625061 Free PMC article.
-
Neuronal hyperactivity-induced oxidant stress promotes in vivo α-synuclein brain spreading.Sci Adv. 2022 Sep 2;8(35):eabn0356. doi: 10.1126/sciadv.abn0356. Epub 2022 Aug 31. Sci Adv. 2022. PMID: 36044566 Free PMC article.
-
Upstream deregulation of calcium signaling in Parkinson's disease.Front Mol Neurosci. 2014 Jun 17;7:53. doi: 10.3389/fnmol.2014.00053. eCollection 2014. Front Mol Neurosci. 2014. PMID: 24987329 Free PMC article. Review.
-
Calcium Channel Antagonists as Disease-Modifying Therapy for Parkinson's Disease: Therapeutic Rationale and Current Status.CNS Drugs. 2016 Dec;30(12):1127-1135. doi: 10.1007/s40263-016-0393-9. CNS Drugs. 2016. PMID: 27826740 Review.
-
Calcium CaV1 channel subtype mRNA expression in Parkinson's disease examined by in situ hybridization.J Mol Neurosci. 2015 Mar;55(3):715-24. doi: 10.1007/s12031-014-0410-8. Epub 2014 Aug 31. J Mol Neurosci. 2015. PMID: 25173401
References
-
- de Lau LM, Breteler MM. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006;5:525–535. - PubMed
-
- Biskup S, Moore DJ. Detrimental deletions: mitochondria, aging and Parkinson’s disease. Bioessays. 2006;28:963–967. - PubMed
-
- Lees AJ, Hardy J, Revesz T. Parkinson’s disease. Lancet. 2009;373:2055–2066. - PubMed
-
- Schapira AH. Mitochondria in the aetiology and pathogenesis of Parkinson’s disease. Lancet Neurol. 2008;7:97–109. - PubMed
-
- Albin RL, Young AB, Penney JB. The functional anatomy of disorders of the basal ganglia. Trends Neurosci. 1995;18:63–64. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
