

Integrative genome analyses identify key somatic driver mutations of small-cell lung cancer
- PMID: 22941188
- PMCID: PMC4915822
- DOI: 10.1038/ng.2396
Integrative genome analyses identify key somatic driver mutations of small-cell lung cancer
Abstract
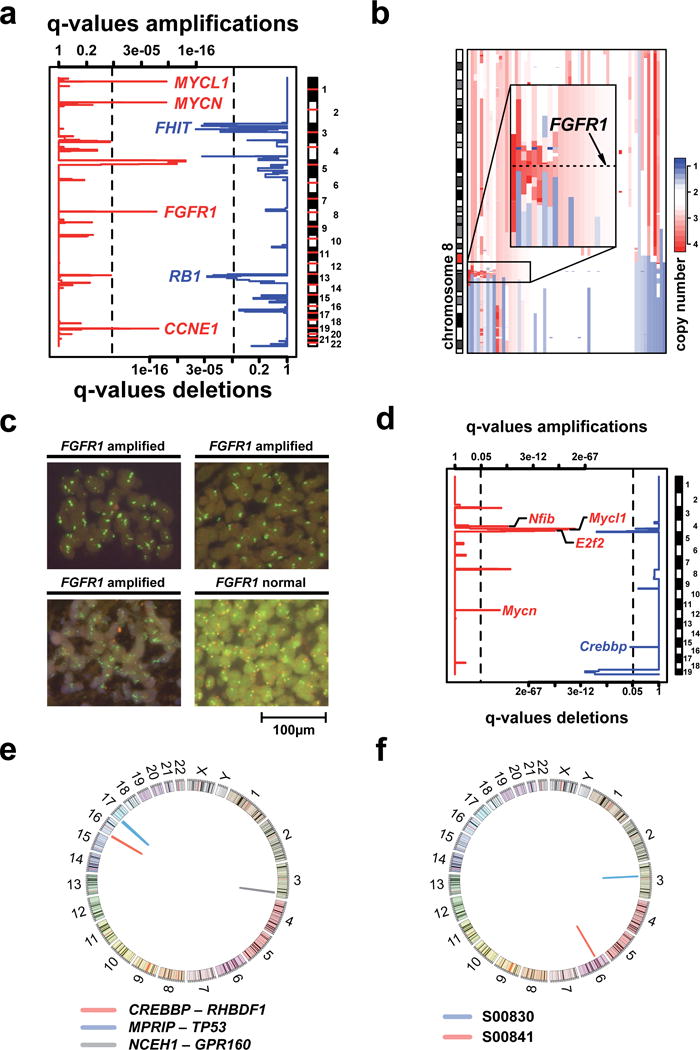
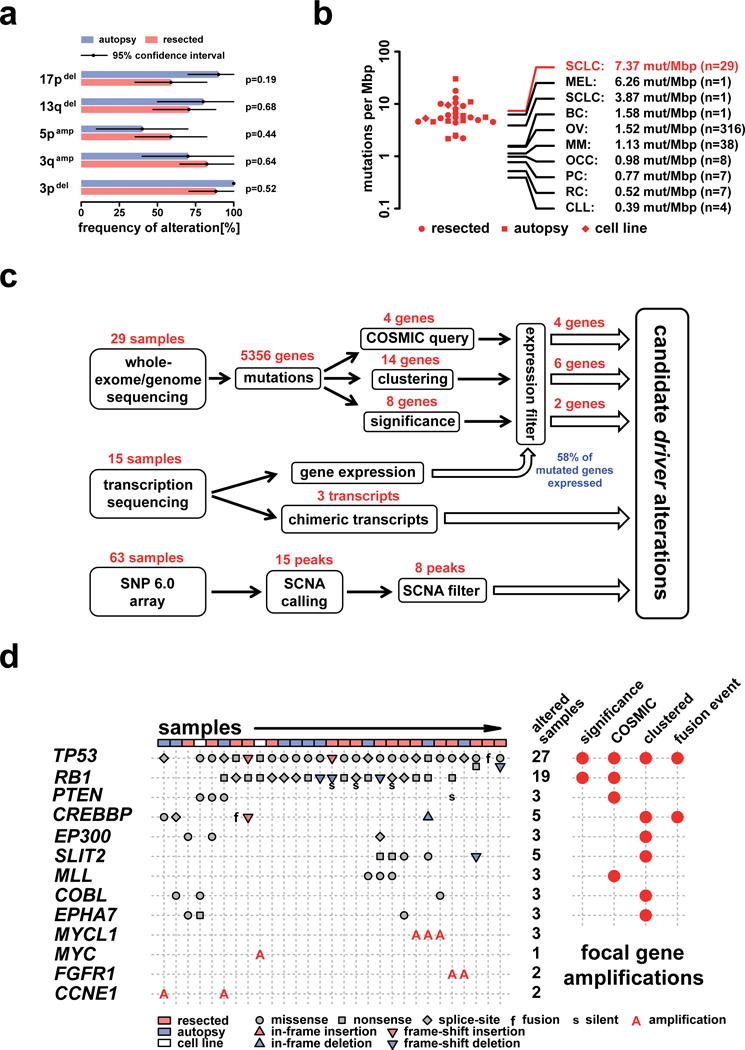
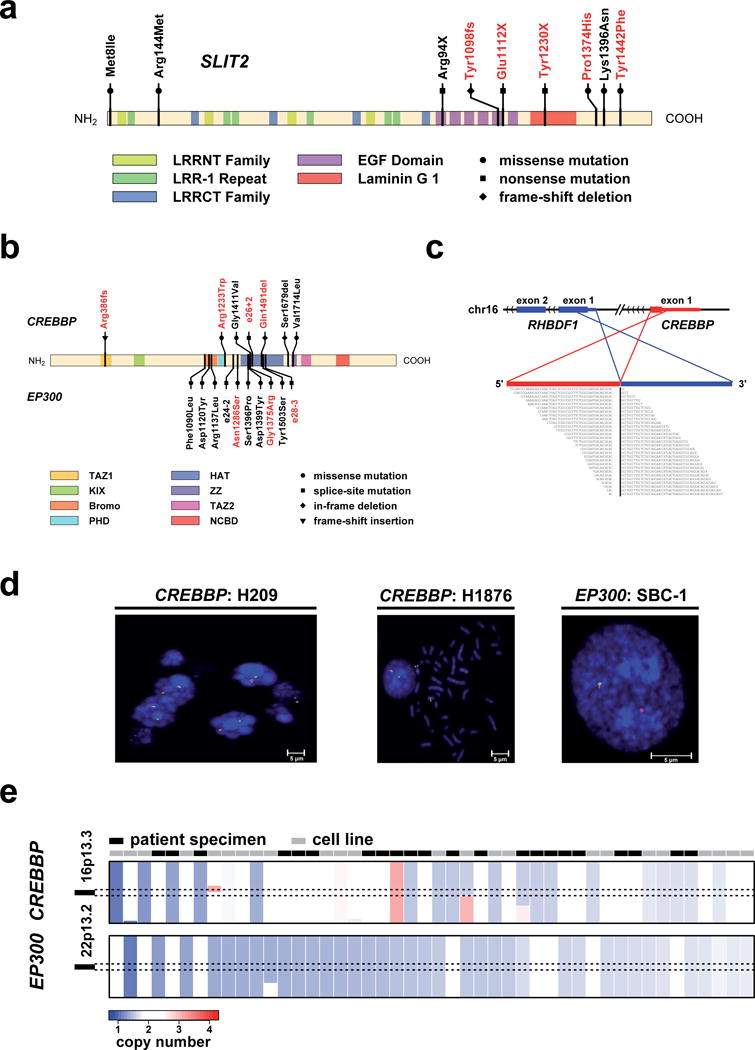
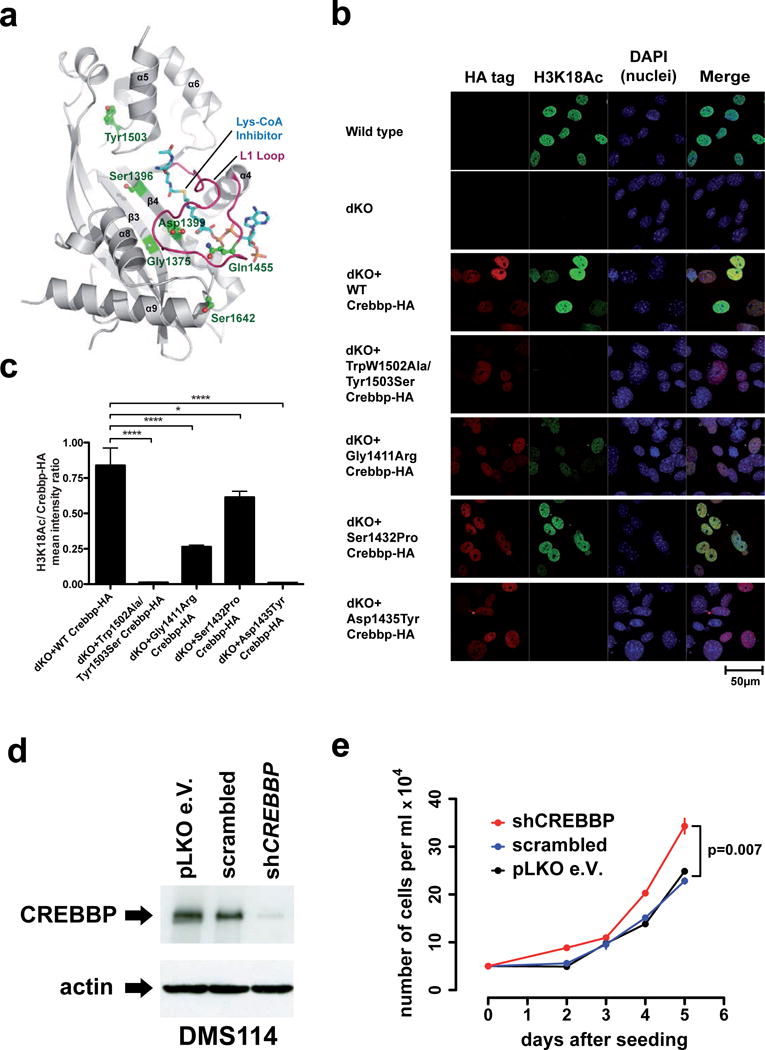
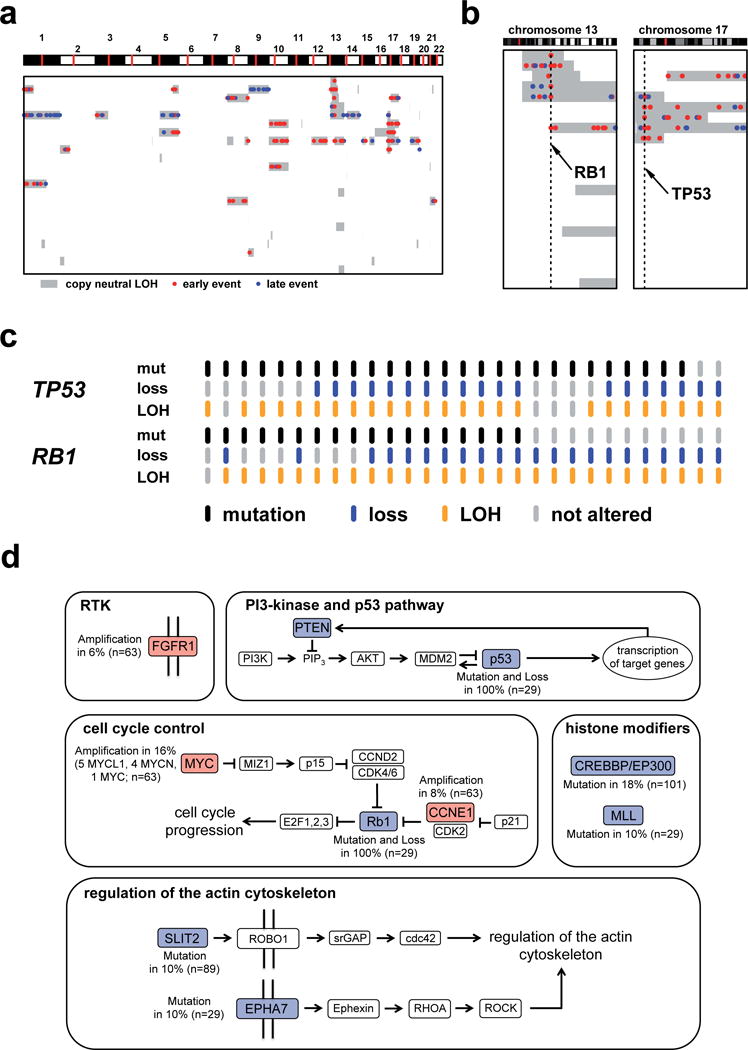
Small-cell lung cancer (SCLC) is an aggressive lung tumor subtype with poor prognosis. We sequenced 29 SCLC exomes, 2 genomes and 15 transcriptomes and found an extremely high mutation rate of 7.4±1 protein-changing mutations per million base pairs. Therefore, we conducted integrated analyses of the various data sets to identify pathogenetically relevant mutated genes. In all cases, we found evidence for inactivation of TP53 and RB1 and identified recurrent mutations in the CREBBP, EP300 and MLL genes that encode histone modifiers. Furthermore, we observed mutations in PTEN, SLIT2 and EPHA7, as well as focal amplifications of the FGFR1 tyrosine kinase gene. Finally, we detected many of the alterations found in humans in SCLC tumors from Tp53 and Rb1 double knockout mice. Our study implicates histone modification as a major feature of SCLC, reveals potentially therapeutically tractable genomic alterations and provides a generalizable framework for the identification of biologically relevant genes in the context of high mutational background.
Figures
Comment in
-
Bringing the genomic landscape of small-cell lung cancer into focus.Nat Genet. 2012 Oct;44(10):1074-5. doi: 10.1038/ng.2415. Nat Genet. 2012. PMID: 23011222 No abstract available.
References
-
- Gustafsson BI, Kidd M, Chan A, Malfertheiner MV, Modlin IM. Bronchopulmonary neuroendocrine tumors. Cancer. 2008;113:5–21. - PubMed
-
- Travis WD. Lung tumours with neuroendocrine differentiation. European journal of cancer. 2009;45(Suppl 1):251–66. - PubMed
-
- van Meerbeeck JP, Fennell DA, De Ruysscher DK. Small-cell lung cancer. Lancet. 2011;378:1741–55. - PubMed
-
- Travis WD, World Health Organization., International Agency for Research on Cancer., International Association for the Study of Lung Cancer & International Academy of Pathology . Pathology and genetics of tumours of the lung, pleura, thymus and heart. IARC Press, Oxford University Press (distributor); Lyon, Oxford: 2004. p. 344.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
