

Beta-catenin-NF-κB interactions in murine hepatocytes: a complex to die for
- PMID: 22941935
- PMCID: PMC3566301
- DOI: 10.1002/hep.26042
Beta-catenin-NF-κB interactions in murine hepatocytes: a complex to die for
Abstract
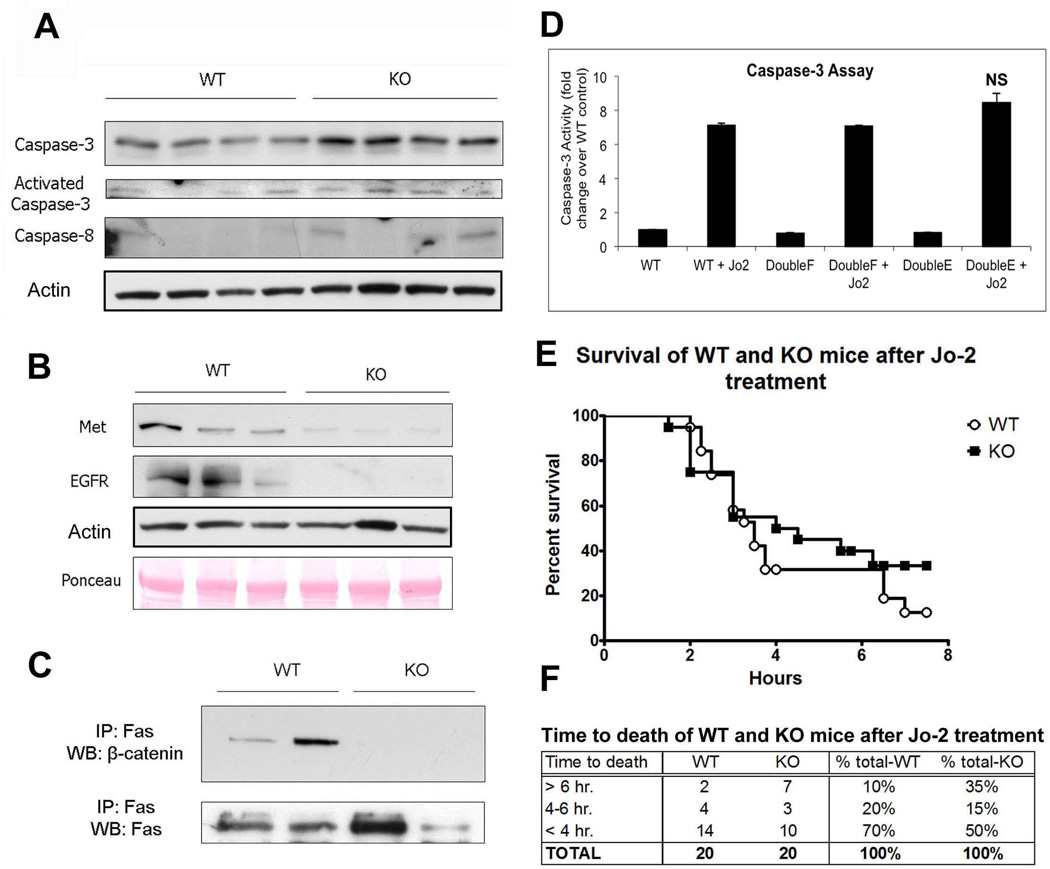
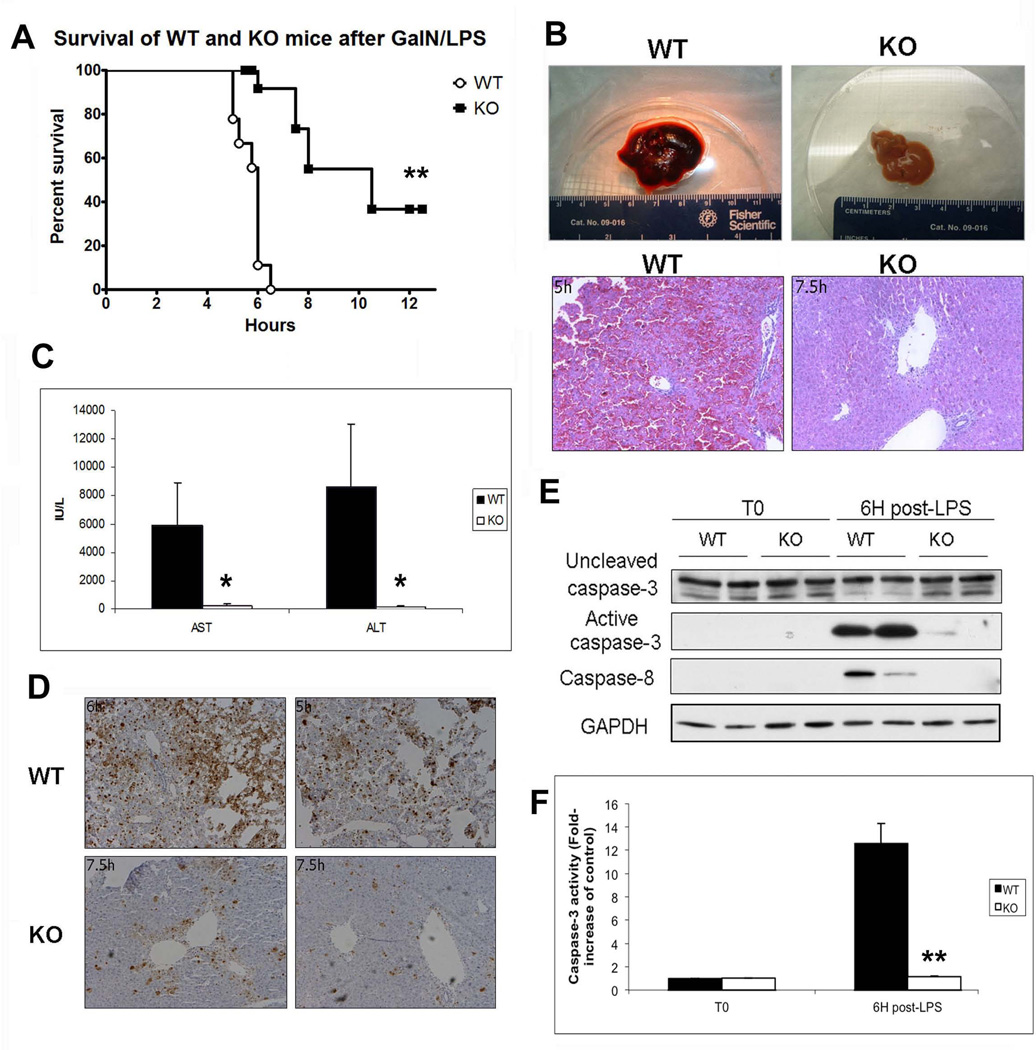
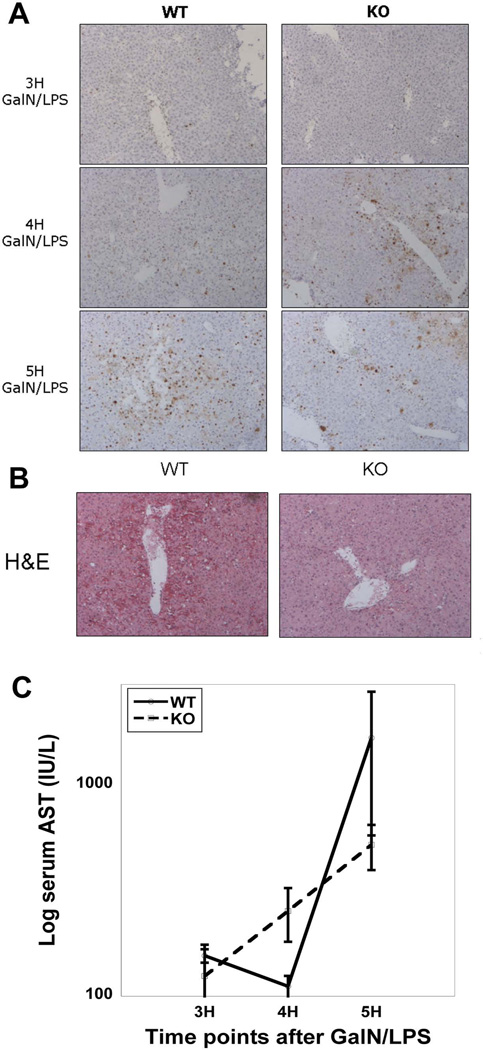
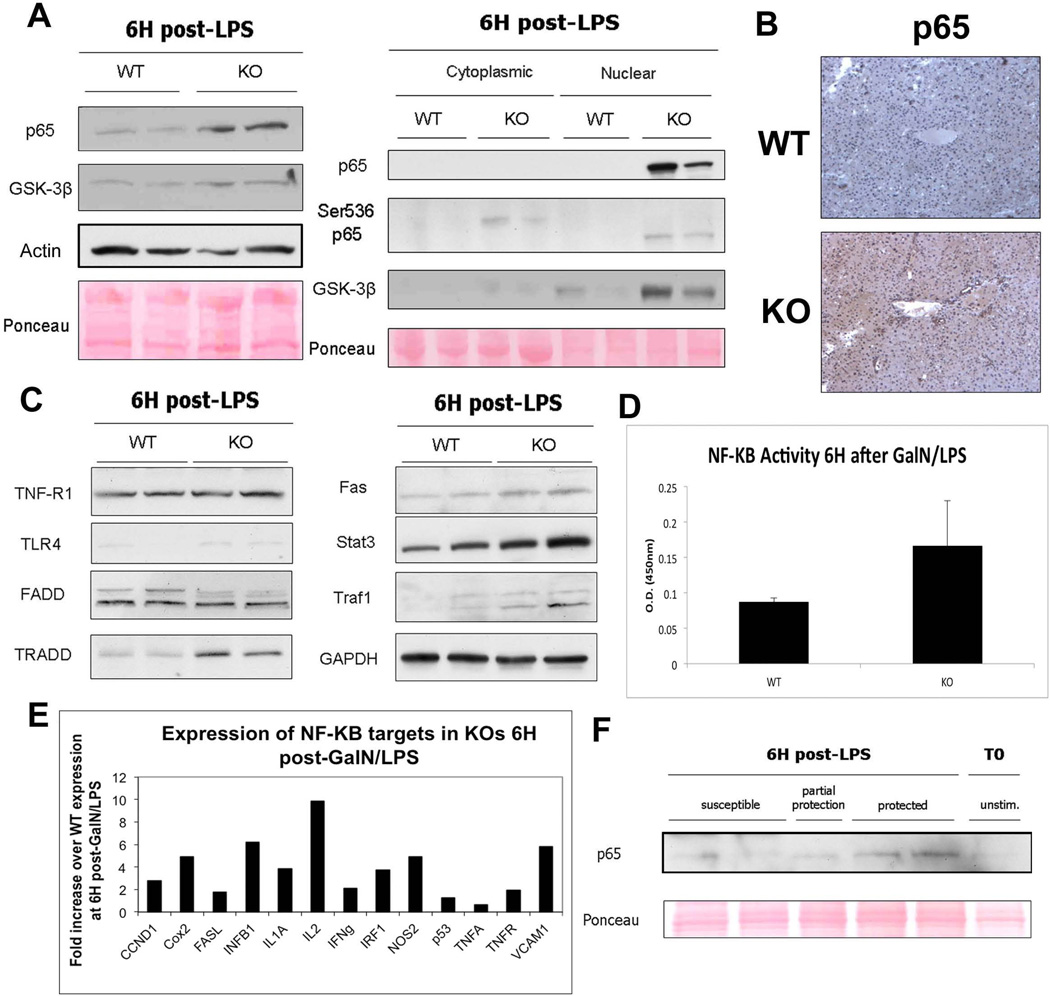
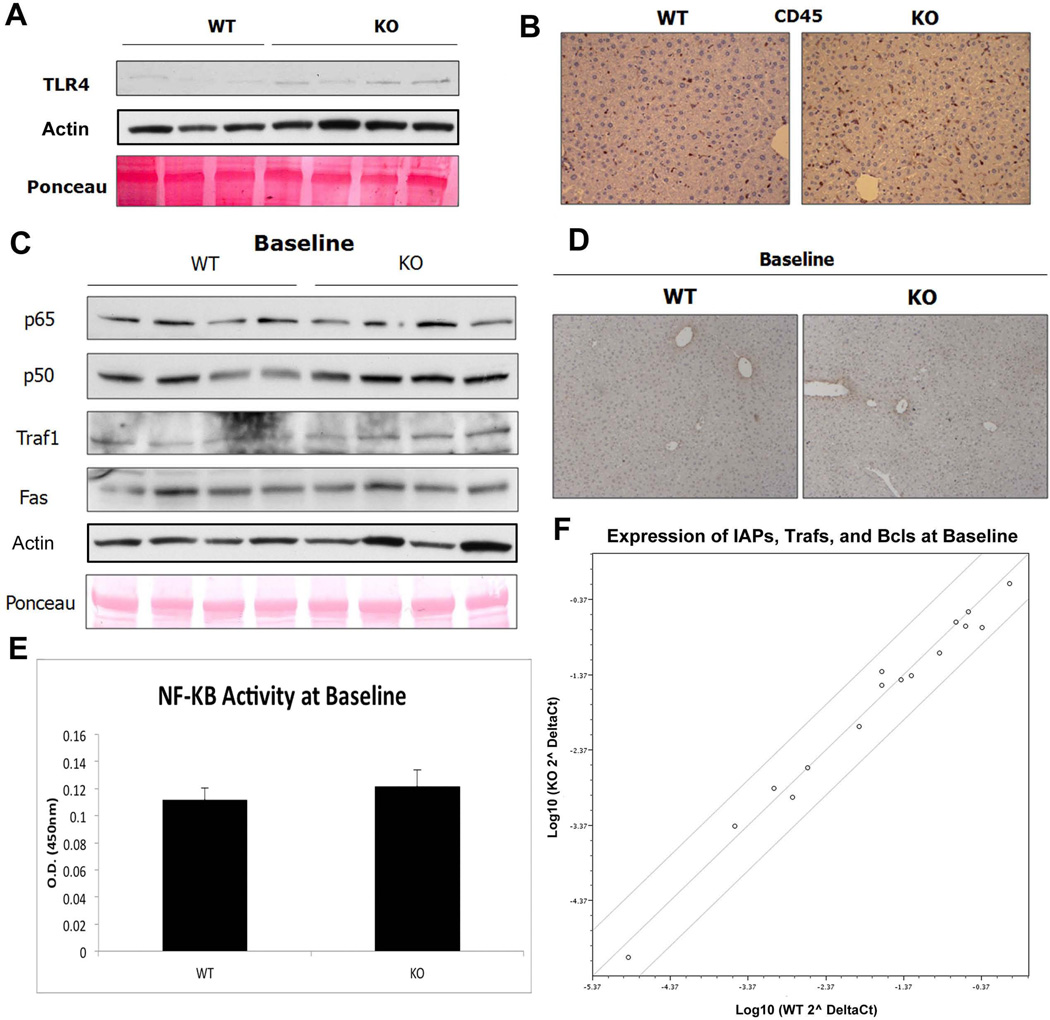
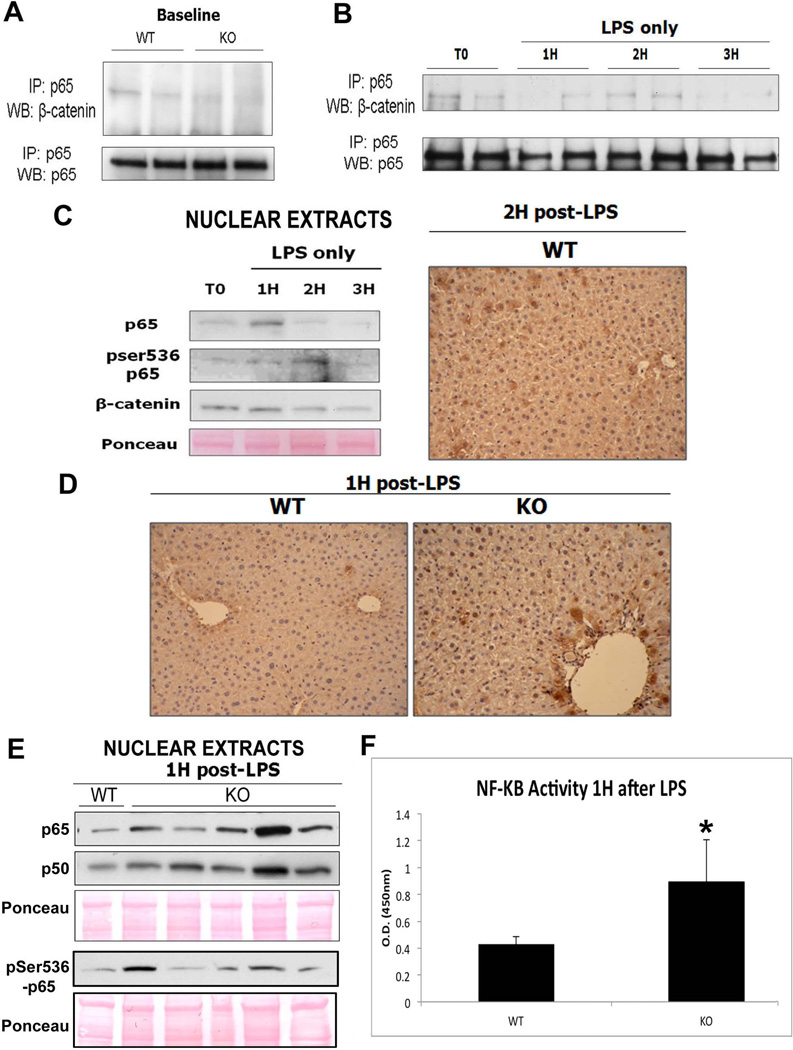
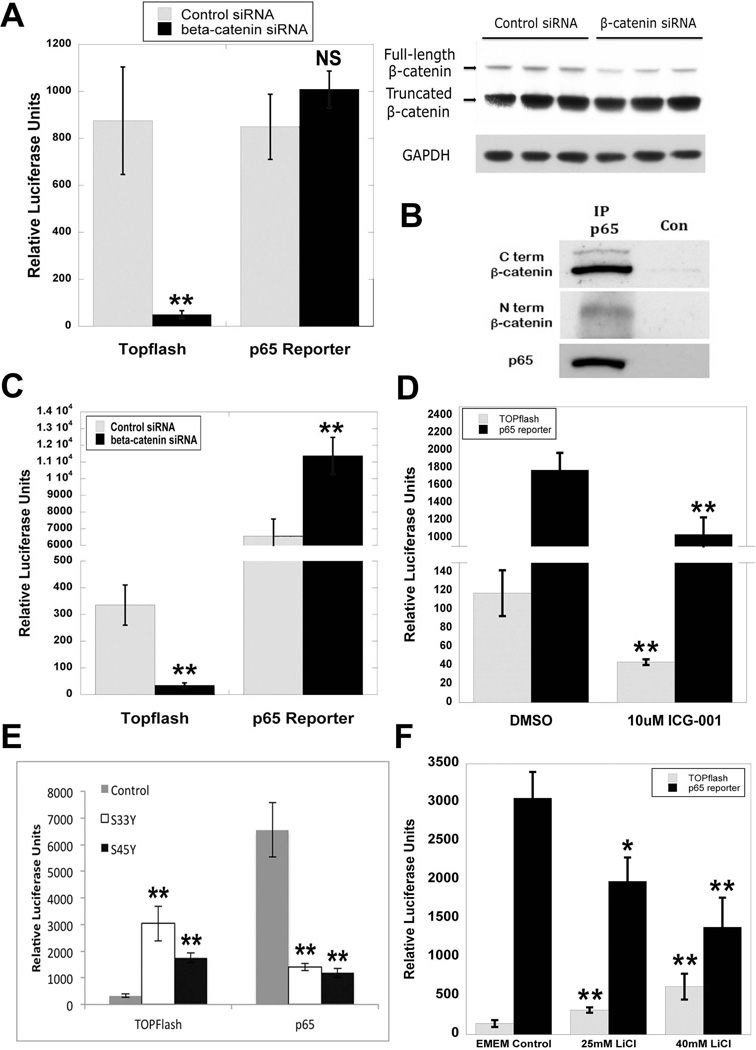
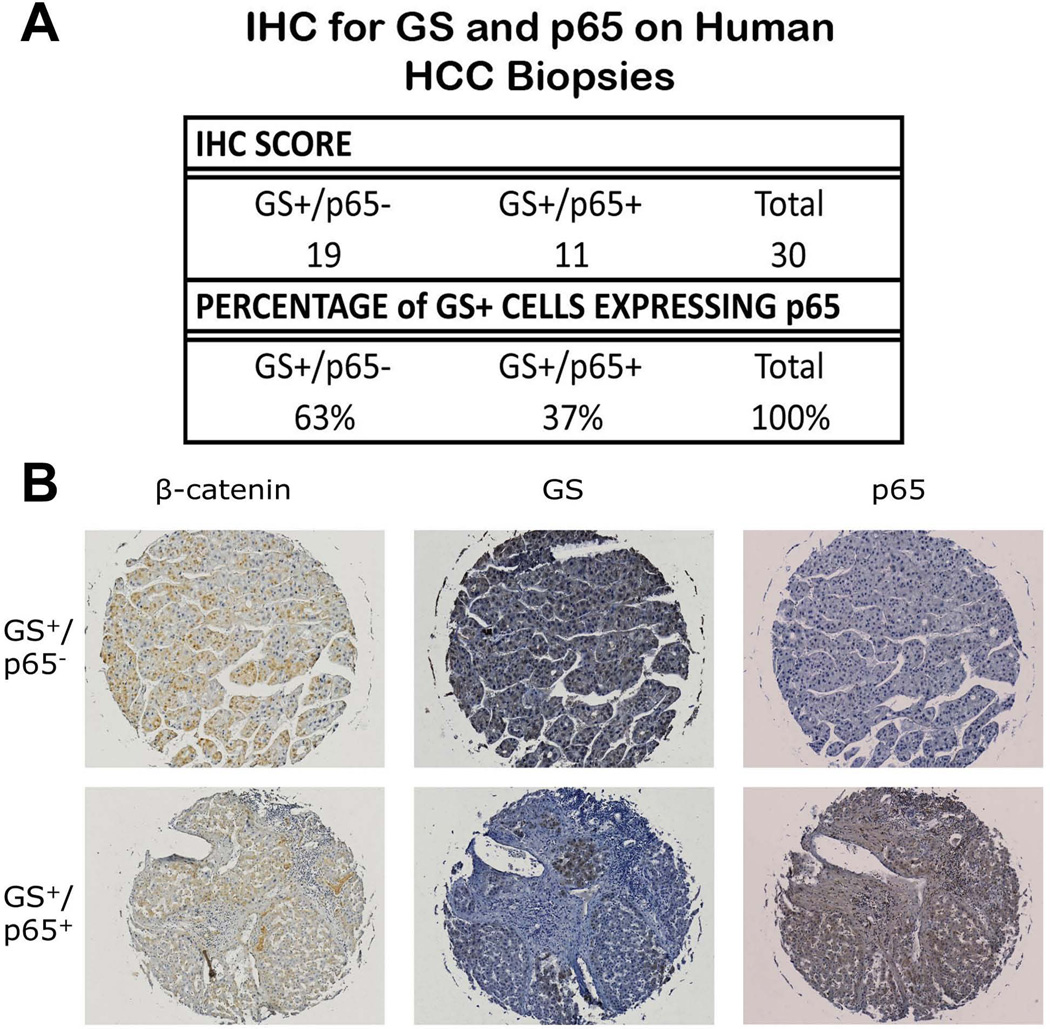
Wnt/β-catenin signaling plays an important role in hepatic homeostasis, especially in liver development, regeneration, and cancer, and loss of β-catenin signaling is often associated with increased apoptosis. To elucidate how β-catenin may be regulating hepatocyte survival, we investigated the susceptibility of β-catenin conditional knockout (KO) mice and their wild-type (WT) littermates to Fas and tumor necrosis factor-α (TNF-α), two common pathways of hepatocyte apoptosis. While comparable detrimental effects from Fas activation were observed in WT and KO, a paradoxical survival benefit was observed in KO mice challenged with D-galactosamine/lipopolysaccharide. KO mice showed significantly lower morbidity and liver injury due to early, robust, and protracted activation of NF-κB in the absence of β-catenin. Enhanced NF-κB activation in KO mice was associated with increased basal inflammation and Toll-like receptor 4 expression and lack of the p65/β-catenin complex in hepatocytes. The p65/β-catenin complex in WT livers underwent temporal dissociation allowing for NF-κB activation to regulate hepatocyte survival following TNF-α-induced hepatic injury. Decrease of total β-catenin protein but not its inactivation induced p65 activity, whereas β-catenin stabilization either chemically or due to mutations repressed it in hepatomas in a dose-dependent manner, whereas β-catenin stabilization repressed it either chemically or due to mutations.
Conclusion: The p65/β-catenin complex in hepatocytes undergoes dynamic changes during TNF-α-induced hepatic injury and plays a critical role in NF-κB activation and cell survival. Modulation of β-catenin levels is a unique mode of regulating NF-κB activity and thus may present novel opportunities in devising therapeutics in specific hepatic injuries.
Copyright © 2012 American Association for the Study of Liver Diseases.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous