

Characterization of the tissue-level Ca2+ signals in spontaneously contracting human myometrium
- PMID: 22947266
- PMCID: PMC4393727
- DOI: 10.1111/j.1582-4934.2012.01626.x
Characterization of the tissue-level Ca2+ signals in spontaneously contracting human myometrium
Abstract
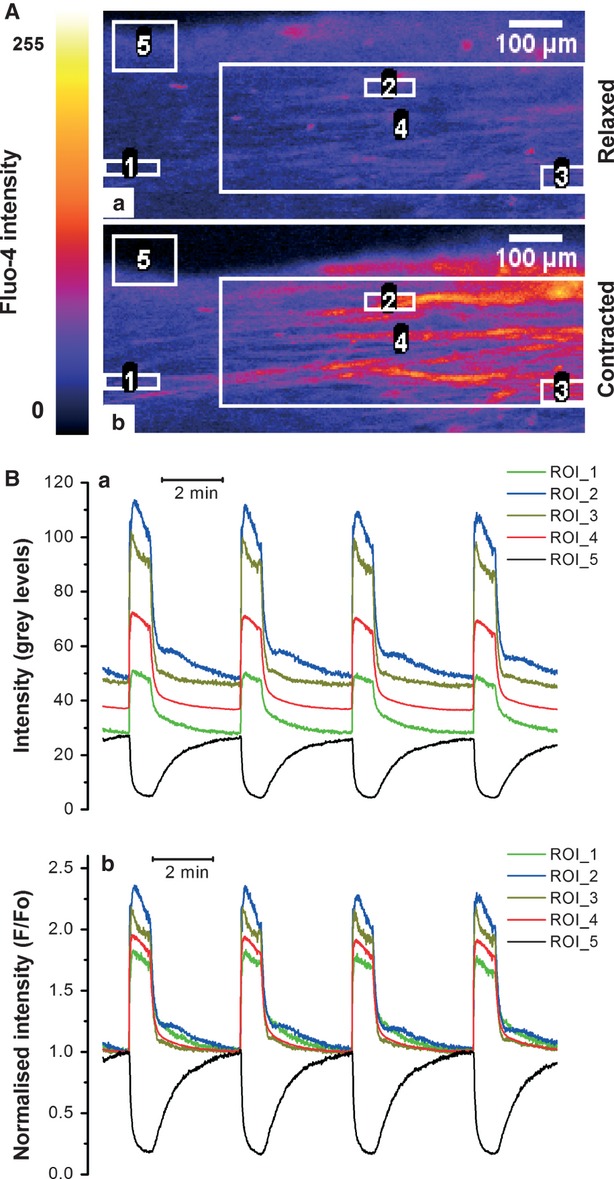
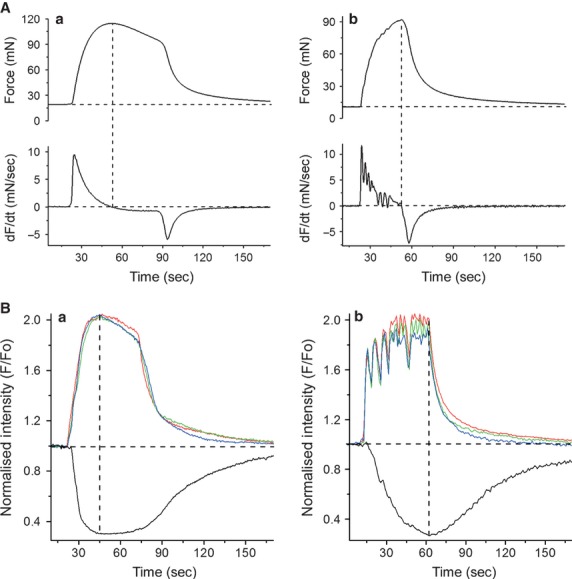
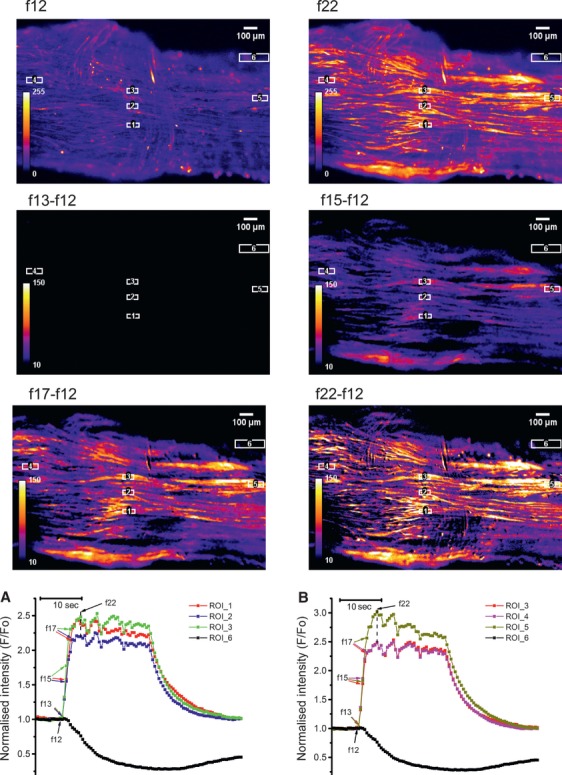
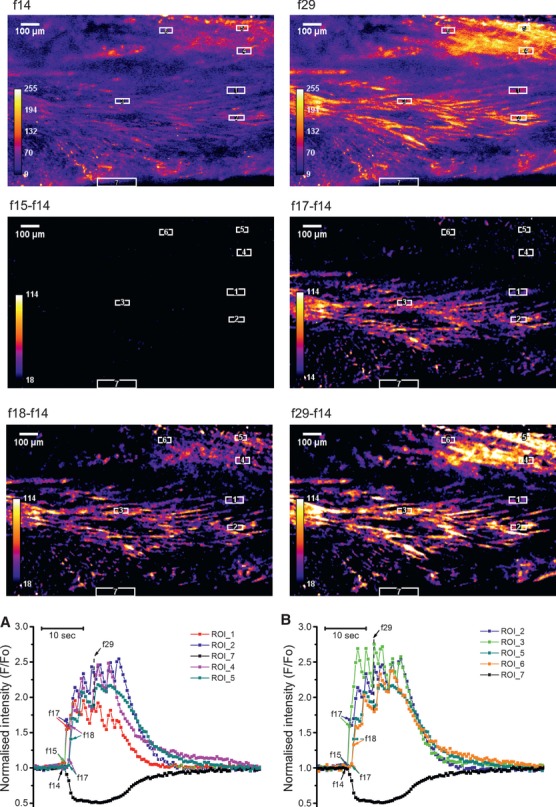
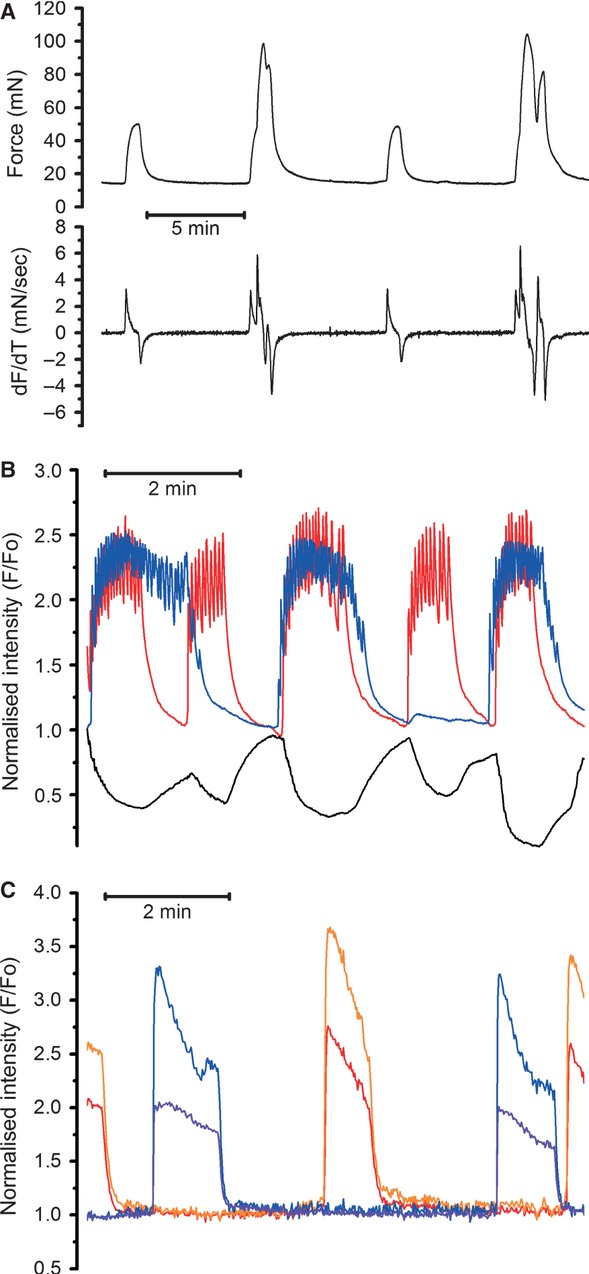
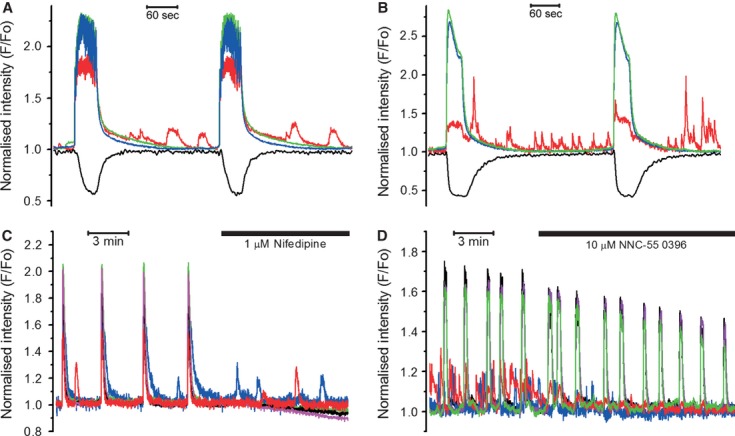
In the labouring uterus, millions of myocytes forming the complex geometrical structure of myometrium contract in synchrony to increase intrauterine pressure, dilate the cervix and eventually expel the foetus through the birth canal. The mechanisms underlying the precise coordination of contractions in human myometrium are not completely understood. In the present study, we have characterized the spatio-temporal properties of tissue-level [Ca(2+)](i) transients in thin slices of intact human myometrium. We found that the waveform of [Ca(2+)](i) transients and isotonic contractions recorded from thin slices was similar to the waveform of isometric contractions recorded from the larger strips in traditional organ bath experiments, suggesting that the spatio-temporal information obtained from thin slices is representative of the whole tissue. By comparing the time course of [Ca(2+)](i) transients in individual cells to that recorded from the bundles of myocytes we found that the majority of myocytes produce rapidly propagating long-lasting [Ca(2+)](i) transients accompanied by contractions. We also found a small number of cells showing desynchronized [Ca(2+)](i) oscillations that did not trigger contractions. The [Ca(2+)](i) oscillations in these cells were insensitive to nifedipine, but readily inhibited by the T-type Ca(2+) channel inhibitor NNC55-0396. In conclusion, our data suggest that the spread of [Ca(2+)](i) signals in human myometrium is achieved via propagation of long-lasting action potentials. The propagation was fast when action potentials propagated along bundles of myocytes and slower when propagating between the bundles of uterine myocytes.
© 2012 The Authors Journal of Cellular and Molecular Medicine © 2012 Foundation for Cellular and Molecular Medicine/Blackwell Publishing Ltd.
Figures
References
-
- Bolton TB. Cholinergic Mechanisms in Smooth-Muscle. Br Med Bull. 1979;35:275–83. - PubMed
-
- Kuriyama H, Kitamura K, Itoh T, et al. Physiological features of visceral smooth muscle cells, with special reference to receptors and ion channels. Physiol Rev. 1998;78:811–920. - PubMed
-
- Wikland M, Lindblom B. Relationship between electrical and mechanical activity of the isolated term-pregnant human myometrium. Eur J Obstet Gynecol Reprod Biol. 1985;20:337–46. - PubMed
-
- Nakao K, Inoue Y, Okabe K, et al. Oxytocin enhances action potentials in pregnant human myometrium - A study with microelectrodes. Am J Obstet Gynecol. 1997;177:222–8. - PubMed
-
- Kawarabayashi T, Izumi H, Ikeda M, et al. Modification by Magnesium of the Excitatory Effect of Oxytocin in Electrical and Mechanical Activities of Pregnant Human Myometrium. Obst Gynecol. 1990;76:183–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
