

Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements
- PMID: 22947866
- PMCID: PMC3414898
- DOI: 10.1016/j.bpj.2012.06.031
Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements
Abstract
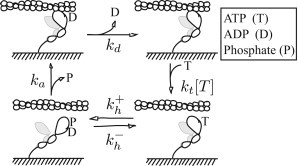
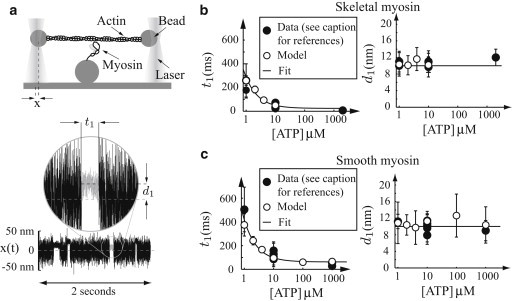
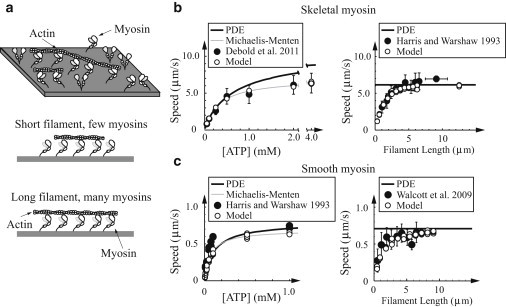
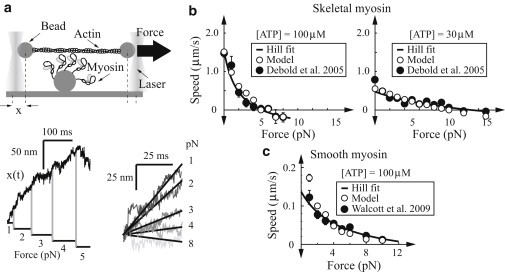
In contracting muscle, individual myosin molecules function as part of a large ensemble, hydrolyzing ATP to power the relative sliding of actin filaments. The technological advances that have enabled direct observation and manipulation of single molecules, including recent experiments that have explored myosin's force-dependent properties, provide detailed insight into the kinetics of myosin's mechanochemical interaction with actin. However, it has been difficult to reconcile these single-molecule observations with the behavior of myosin in an ensemble. Here, using a combination of simulations and theory, we show that the kinetic mechanism derived from single-molecule experiments describes ensemble behavior; but the connection between single molecule and ensemble is complex. In particular, even in the absence of external force, internal forces generated between myosin molecules in a large ensemble accelerate ADP release and increase how far actin moves during a single myosin attachment. These myosin-induced changes in strong binding lifetime and attachment distance cause measurable properties, such as actin speed in the motility assay, to vary depending on the number of myosin molecules interacting with an actin filament. This ensemble-size effect challenges the simple detachment limited model of motility, because even when motility speed is limited by ADP release, increasing attachment rate can increase motility speed.
Copyright © 2012 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Lymn R.W., Taylor E.W. Mechanism of adenosine triphosphate hydrolysis by actomyosin. Biochemistry. 1971;10:4617–4624. - PubMed
-
- Finer J.T., Simmons R.M., Spudich J.A. Single myosin molecule mechanics: picoNewton forces and nanometer steps. Nature. 1994;368:113–119. - PubMed
-
- Ishijima A., Harada Y., Yanagida T. Single-molecule analysis of the actomyosin motor using nano-manipulation. Biochem. Biophys. Res. Commun. 1994;199:1057–1063. - PubMed
-
- Molloy J.E., Burns J.E., White D.C. Movement and force produced by a single myosin head. Nature. 1995;378:209–212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
