

HtrA1 is a novel antagonist controlling fibroblast growth factor (FGF) signaling via cleavage of FGF8
- PMID: 22949504
- PMCID: PMC3486155
- DOI: 10.1128/MCB.00872-12
HtrA1 is a novel antagonist controlling fibroblast growth factor (FGF) signaling via cleavage of FGF8
Abstract
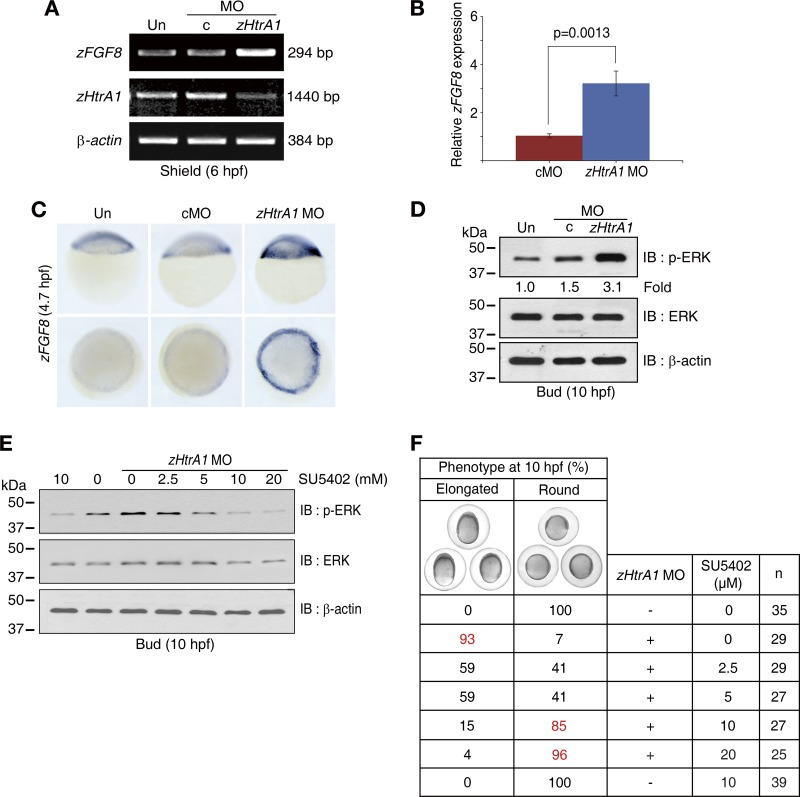
Accumulating evidence suggests that HtrA1 (high-temperature requirement A1) is involved in modulating crucial cellular processes and implicated in life-threatening diseases, such as cancer and neuropathological disorders; however, the exact functions of this protease in vivo remain unknown. Here, we show that loss of HtrA1 function increases fibroblast growth factor 8 (FGF8) mRNA levels and triggers activation of FGF signaling, resulting in dorsalization in zebrafish embryos. Notably, HtrA1 directly cleaves FGF8 in the extracellular region, and this cleavage results in decreased activation of FGF signaling, which is essential for many physiological processes. Therefore, HtrA1 is indispensable for dorsoventral patterning in early zebrafish embryogenesis and serves as a key upstream regulator of FGF signaling through the control of FGF levels. Furthermore, this study offers insight into new strategies to control human diseases associated with HtrA1 and FGF signaling.
Figures





Similar articles
-
SCUBE3 (signal peptide-CUB-EGF domain-containing protein 3) modulates fibroblast growth factor signaling during fast muscle development.J Biol Chem. 2014 Jul 4;289(27):18928-42. doi: 10.1074/jbc.M114.551929. Epub 2014 May 21. J Biol Chem. 2014. PMID: 24849601 Free PMC article.
-
Fgf8 and Fgf3 are required for zebrafish ear placode induction, maintenance and inner ear patterning.Mech Dev. 2002 Nov;119(1):91-108. doi: 10.1016/s0925-4773(02)00343-x. Mech Dev. 2002. PMID: 12385757
-
Maternal control of vertebrate dorsoventral axis formation and epiboly by the POU domain protein Spg/Pou2/Oct4.Development. 2006 Jul;133(14):2757-70. doi: 10.1242/dev.02391. Epub 2006 Jun 14. Development. 2006. PMID: 16775002
-
Roles of FGF8 subfamily in embryogenesis and oral‑maxillofacial diseases (Review).Int J Oncol. 2019 Mar;54(3):797-806. doi: 10.3892/ijo.2019.4677. Epub 2019 Jan 7. Int J Oncol. 2019. PMID: 30628659 Review.
-
Numerous isoforms of Fgf8 reflect its multiple roles in the developing brain.J Cell Physiol. 2011 Jul;226(7):1722-6. doi: 10.1002/jcp.22587. J Cell Physiol. 2011. PMID: 21506104 Free PMC article. Review.
Cited by
-
Effects of Yersinia ruckeri invasion on the proteome of the Chinook salmon cell line CHSE-214.Sci Rep. 2020 Jul 16;10(1):11840. doi: 10.1038/s41598-020-68903-5. Sci Rep. 2020. PMID: 32678312 Free PMC article.
-
Exploring mechanisms of FGF signalling through the lens of structural biology.Nat Rev Mol Cell Biol. 2013 Mar;14(3):166-80. doi: 10.1038/nrm3528. Epub 2013 Feb 13. Nat Rev Mol Cell Biol. 2013. PMID: 23403721 Free PMC article. Review.
-
Serine protease HtrA1 as an inhibitor on proliferation invasion and migration of gastric cancer.Med Oncol. 2015 Apr;32(4):112. doi: 10.1007/s12032-015-0524-z. Epub 2015 Mar 12. Med Oncol. 2015. PMID: 25761858
-
Asian age-related macular degeneration: from basic science research perspective.Eye (Lond). 2019 Jan;33(1):34-49. doi: 10.1038/s41433-018-0225-x. Epub 2018 Oct 12. Eye (Lond). 2019. PMID: 30315261 Free PMC article. Review.
-
Endocytosis of Fgf8 is a double-stage process and regulates spreading and signaling.PLoS One. 2014 Jan 20;9(1):e86373. doi: 10.1371/journal.pone.0086373. eCollection 2014. PLoS One. 2014. PMID: 24466061 Free PMC article.
References
-
- Baldi A, et al. 2002. The HtrA1 serine protease is down-regulated during human melanoma progression and represses growth of metastatic melanoma cells. Oncogene 21:6684–6688 - PubMed
-
- Bottcher RT, Niehrs C. 2005. Fibroblast growth factor signaling during early vertebrate development. Endocr Rev. 26:63–77 - PubMed
-
- Bottcher RT, Pollet N, Delius H, Niehrs C. 2004. The transmembrane protein XFLRT3 forms a complex with FGF receptors and promotes FGF signalling. Nat. Cell Biol. 6:38–44 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases