

Muscle length and myonuclear position are independently regulated by distinct Dynein pathways
- PMID: 22951643
- PMCID: PMC3445310
- DOI: 10.1242/dev.079178
Muscle length and myonuclear position are independently regulated by distinct Dynein pathways
Abstract
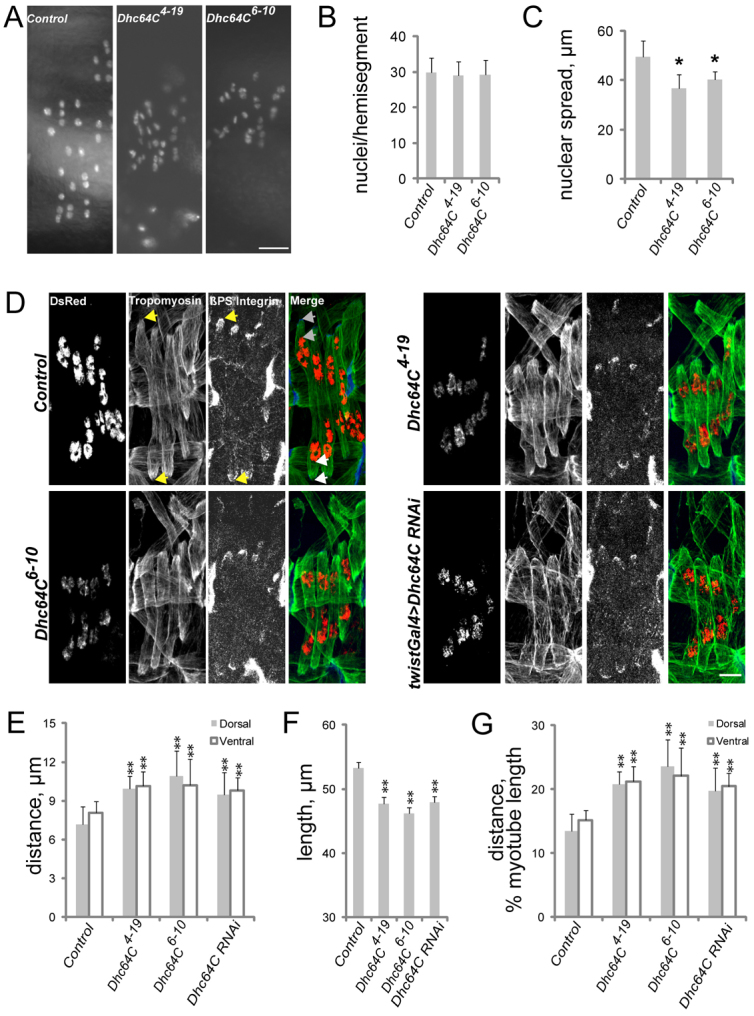
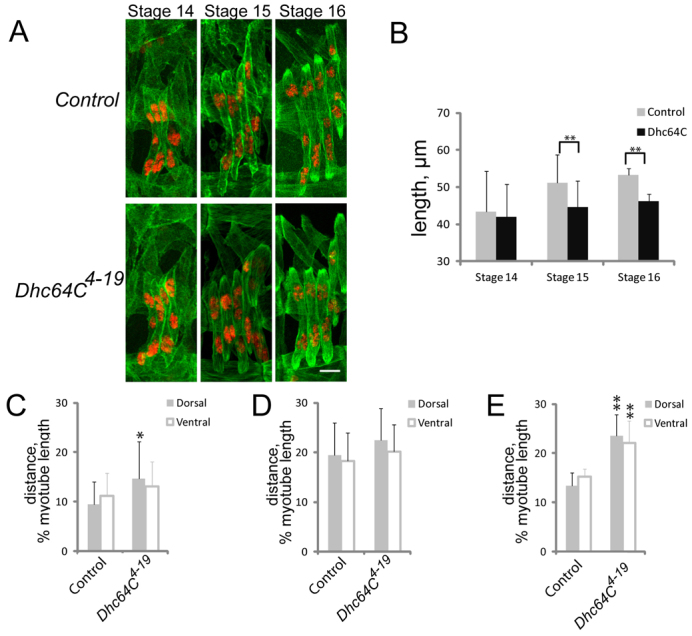
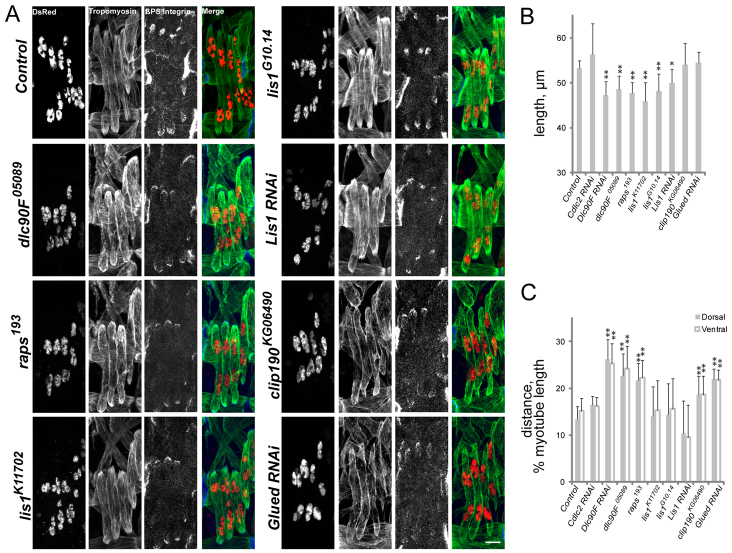
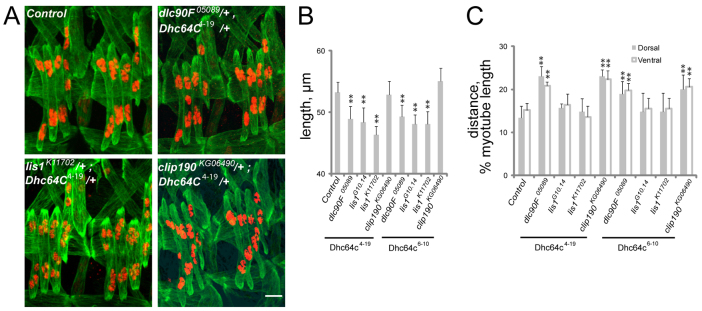
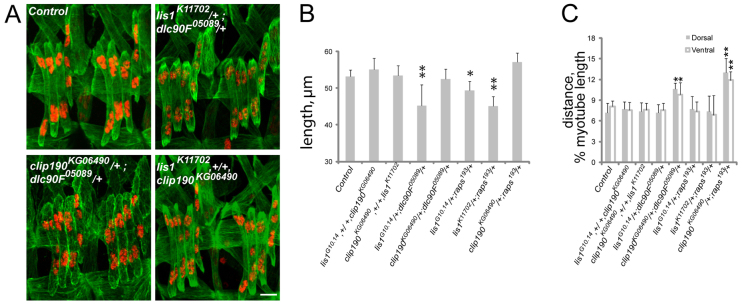
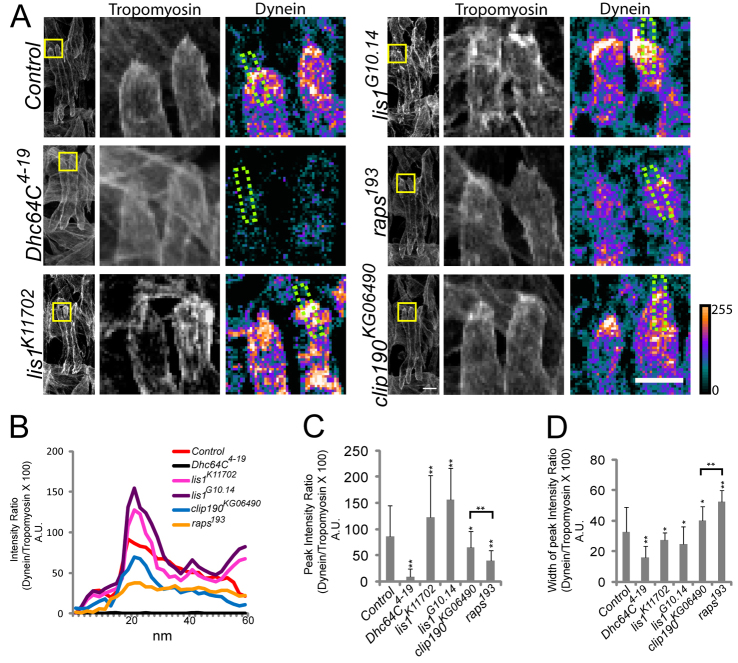
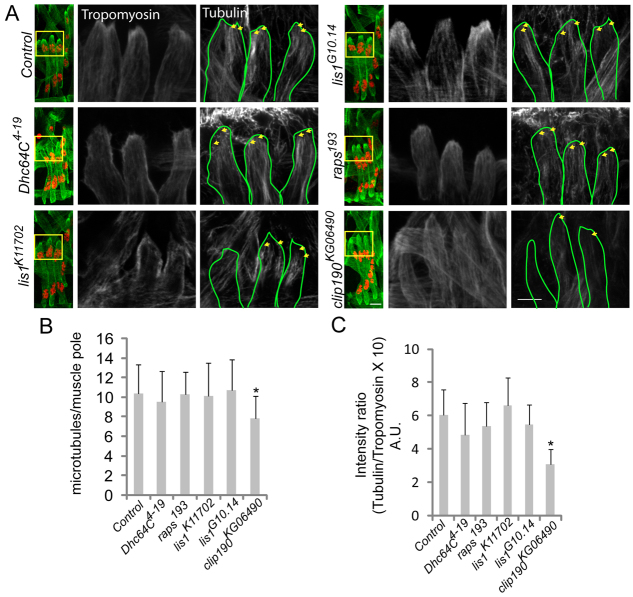
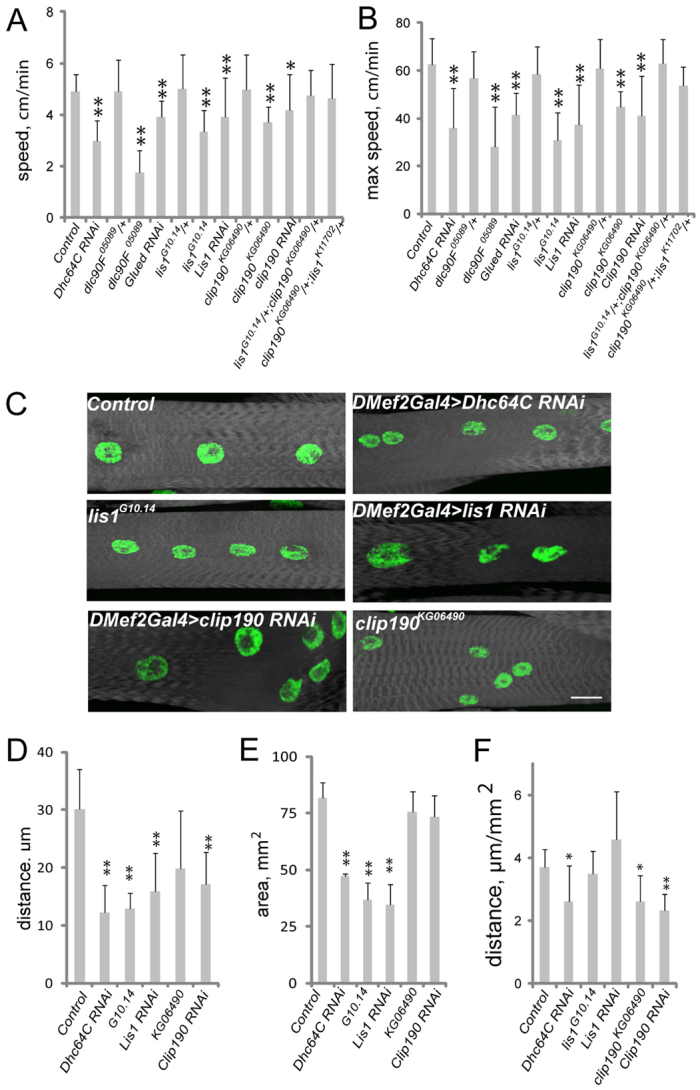
Various muscle diseases present with aberrant muscle cell morphologies characterized by smaller myofibers with mispositioned nuclei. The mechanisms that normally control these processes, whether they are linked, and their contribution to muscle weakness in disease, are not known. We examined the role of Dynein and Dynein-interacting proteins during Drosophila muscle development and found that several factors, including Dynein heavy chain, Dynein light chain and Partner of inscuteable, contribute to the regulation of both muscle length and myonuclear positioning. However, Lis1 contributes only to Dynein-dependent muscle length determination, whereas CLIP-190 and Glued contribute only to Dynein-dependent myonuclear positioning. Mechanistically, microtubule density at muscle poles is decreased in CLIP-190 mutants, suggesting that microtubule-cortex interactions facilitate myonuclear positioning. In Lis1 mutants, Dynein hyperaccumulates at the muscle poles with a sharper localization pattern, suggesting that retrograde trafficking contributes to muscle length. Both Lis1 and CLIP-190 act downstream of Dynein accumulation at the cortex, suggesting that they specify Dynein function within a single location. Finally, defects in muscle length or myonuclear positioning correlate with impaired muscle function in vivo, suggesting that both processes are essential for muscle function.
Figures
References
-
- Bate M. (1990). The embryonic development of larval muscles in Drosophila. Development 110, 791-804 - PubMed
-
- Caggese C., Moschetti R., Ragone G., Barsanti P., Caizzi R. (2001). dtctex-1, the Drosophila melanogaster homolog of a putative murine t-complex distorter encoding a dynein light chain, is required for production of functional sperm. Mol. Genet. Genomics 265, 436-444 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
