

Hallmarks of processivity in glycoside hydrolases from crystallographic and computational studies of the Serratia marcescens chitinases
- PMID: 22952223
- PMCID: PMC3476299
- DOI: 10.1074/jbc.M112.402149
Hallmarks of processivity in glycoside hydrolases from crystallographic and computational studies of the Serratia marcescens chitinases
Abstract
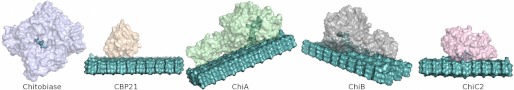
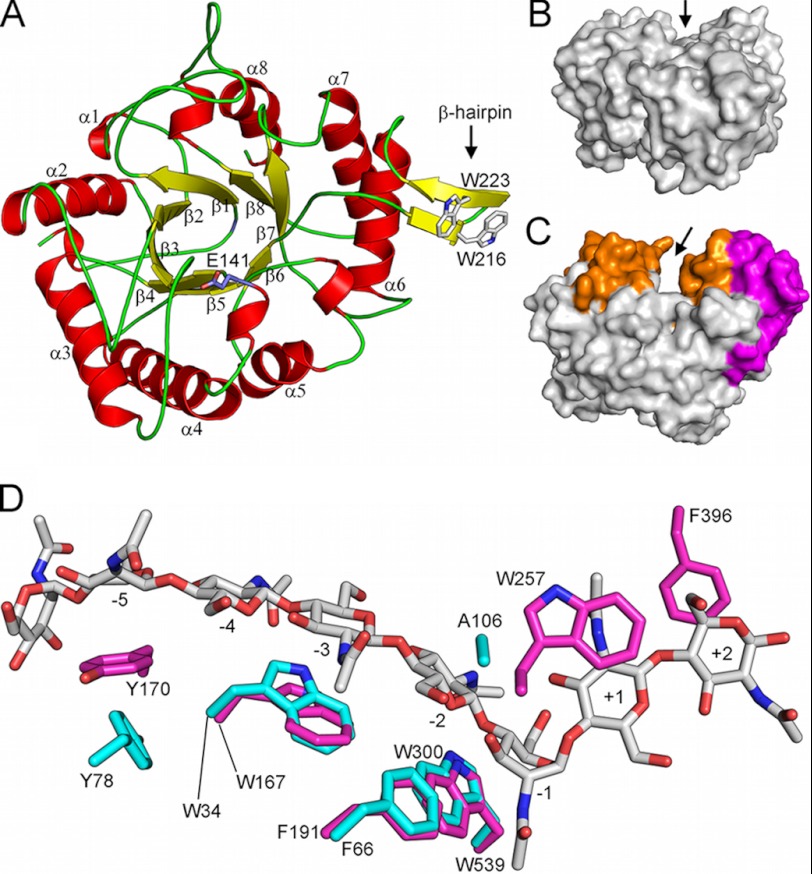
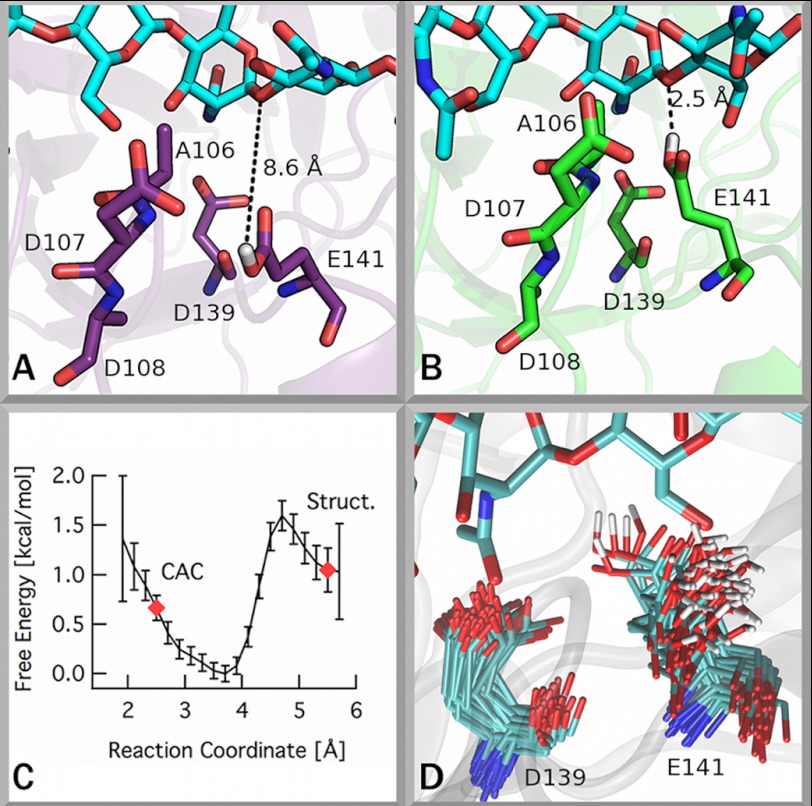
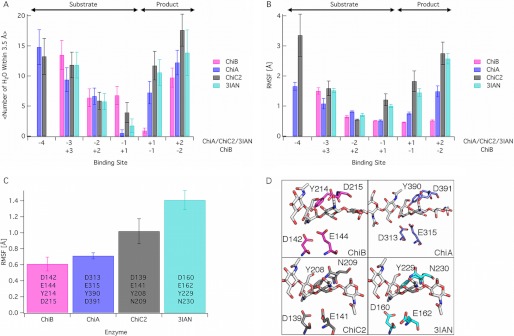
Degradation of recalcitrant polysaccharides in nature is typically accomplished by mixtures of processive and nonprocessive glycoside hydrolases (GHs), which exhibit synergistic activity wherein nonprocessive enzymes provide new sites for productive attachment of processive enzymes. GH processivity is typically attributed to active site geometry, but previous work has demonstrated that processivity can be tuned by point mutations or removal of single loops. To gain additional insights into the differences between processive and nonprocessive enzymes that give rise to their synergistic activities, this study reports the crystal structure of the catalytic domain of the GH family 18 nonprocessive endochitinase, ChiC, from Serratia marcescens. This completes the structural characterization of the co-evolved chitinolytic enzymes from this bacterium and enables structural analysis of their complementary functions. The ChiC catalytic module reveals a shallow substrate-binding cleft that lacks aromatic residues vital for processivity, a calcium-binding site not previously seen in GH18 chitinases, and, importantly, a displaced catalytic acid (Glu-141), suggesting flexibility in the catalytic center. Molecular dynamics simulations of two processive chitinases (ChiA and ChiB), the ChiC catalytic module, and an endochitinase from Lactococcus lactis show that the nonprocessive enzymes have more flexible catalytic machineries and that their bound ligands are more solvated and flexible. These three features, which relate to the more dynamic on-off ligand binding processes associated with nonprocessive action, correlate to experimentally measured differences in processivity of the S. marcescens chitinases. These newly defined hallmarks thus appear to be key dynamic metrics in determining processivity in GH enzymes complementing structural insights.
Figures
References
-
- Himmel M. E., Ding S. Y., Johnson D. K., Adney W. S., Nimlos M. R., Brady J. W., Foust T. D. (2007) Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science 315, 804–807 - PubMed
-
- Vaaje-Kolstad G., Westereng B., Horn S. J., Liu Z., Zhai H., Sørlie M., Eijsink V. G. (2010) An oxidative enzyme boosting the enzymatic conversion of recalcitrant polysaccharides. Science 330, 219–222 - PubMed
-
- Mba Medie F., Davies G. J., Drancourt M., Henrissat B. (2012) Genome analyses highlight the different biological roles of cellulases. Nat. Rev. Microbiol. 10, 227–234 - PubMed
-
- Bayer E. A., Belaich J. P., Shoham Y., Lamed R. (2004) The cellulosomes: multienzyme machines for degradation of plant cell wall polysaccharides. Annu. Rev. Microbiol. 58, 521–554 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
