

Bromodomain and extra-terminal (BET) bromodomain inhibition activate transcription via transient release of positive transcription elongation factor b (P-TEFb) from 7SK small nuclear ribonucleoprotein
- PMID: 22952229
- PMCID: PMC3476326
- DOI: 10.1074/jbc.M112.410746
Bromodomain and extra-terminal (BET) bromodomain inhibition activate transcription via transient release of positive transcription elongation factor b (P-TEFb) from 7SK small nuclear ribonucleoprotein
Erratum in
- J Biol Chem. 2013 Apr 26;288(17):12214
Abstract
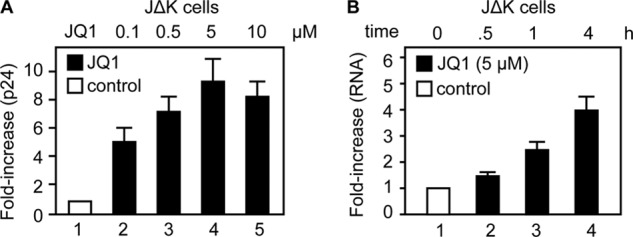
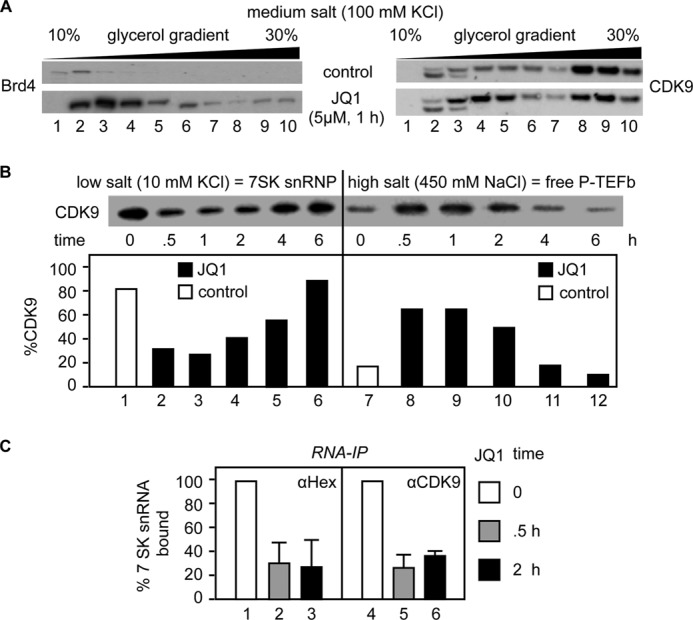
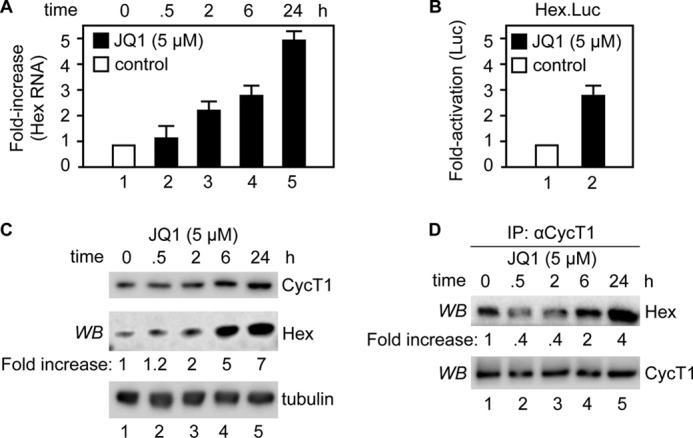
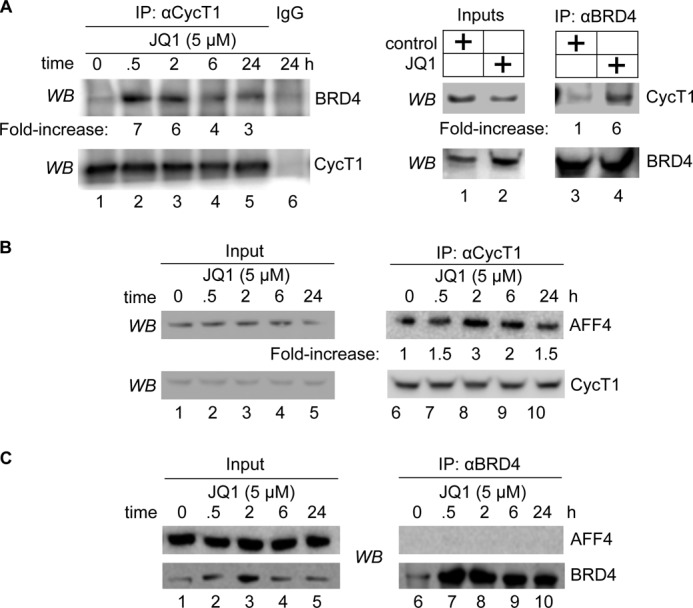
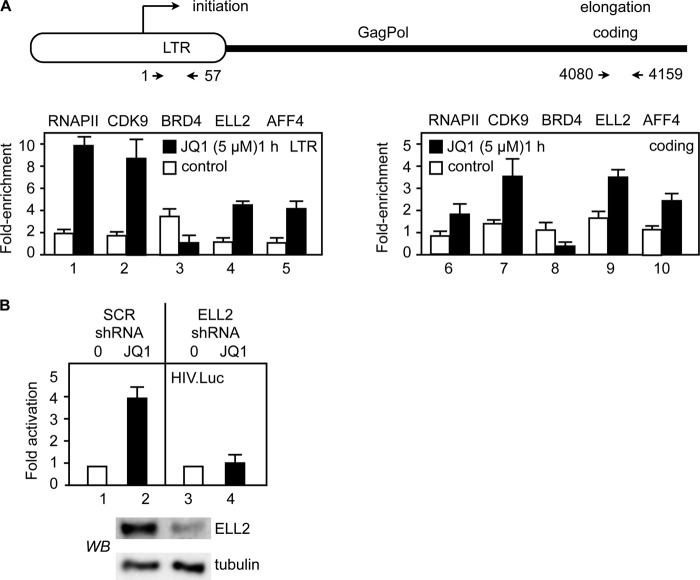
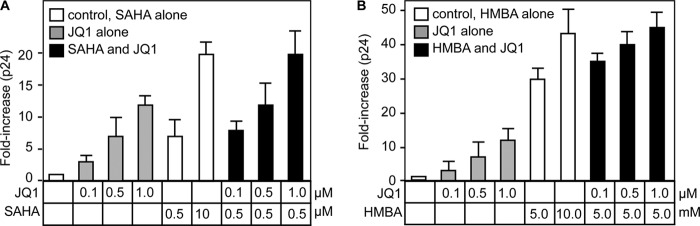
By phosphorylating elongation factors and the C-terminal domain of RNA polymerase II, the positive transcription elongation factor b (P-TEFb) is the critical kinase for transcription elongation and co-transcriptional processing of eukaryotic genes. It exists in inactive small nuclear ribonucleoprotein (7SK snRNP) and active (free P-TEFb) complexes in cells. The P-TEFb equilibrium determines the state of cellular activation, proliferation, and differentiation. Free P-TEFb, which is required for growth, can be recruited to RNA polymerase II via transcription factors, BRD4, or the super elongation complex (SEC). UV light, various signaling cascades, transcriptional blockade, or compounds such as hexamethylene bisacetamide (HMBA), suberoylanilide hydroxamic acid (SAHA), and other histone deacetylase inhibitors lead to a rapid release of free P-TEFb, followed by its reassembly into the 7SK snRNP. As a consequence, transcription of HEXIM1, a critical 7SK snRNP subunit, and HIV is induced. In this study, we found that a bromodomain and extra-terminal (BET) bromodomain inhibitor, JQ1, which inhibits BRD4 by blocking its association with chromatin, also leads to the rapid release of free P-TEFb from the 7SK snRNP. Indeed, JQ1 transiently increased levels of free P-TEFb and BRD4·P-TEFb and SEC·P-TEFb complexes in cells. As a consequence, the levels of HEXIM1 and HIV proteins rose. Importantly, the knockdown of ELL2, a subunit of the SEC, blocked the ability of JQ1 to increase HIV transcription. Finally, the effects of JQ1 and HMBA or SAHA on the P-TEFb equilibrium were cooperative. We conclude that HMBA, SAHA, and JQ1 affect transcription elongation by a similar and convergent mechanism.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
