

Transcriptome and proteome dynamics of a light-dark synchronized bacterial cell cycle
- PMID: 22952681
- PMCID: PMC3430701
- DOI: 10.1371/journal.pone.0043432
Transcriptome and proteome dynamics of a light-dark synchronized bacterial cell cycle
Abstract
Background: Growth of the ocean's most abundant primary producer, the cyanobacterium Prochlorococcus, is tightly synchronized to the natural 24-hour light-dark cycle. We sought to quantify the relationship between transcriptome and proteome dynamics that underlie this obligate photoautotroph's highly choreographed response to the daily oscillation in energy supply.
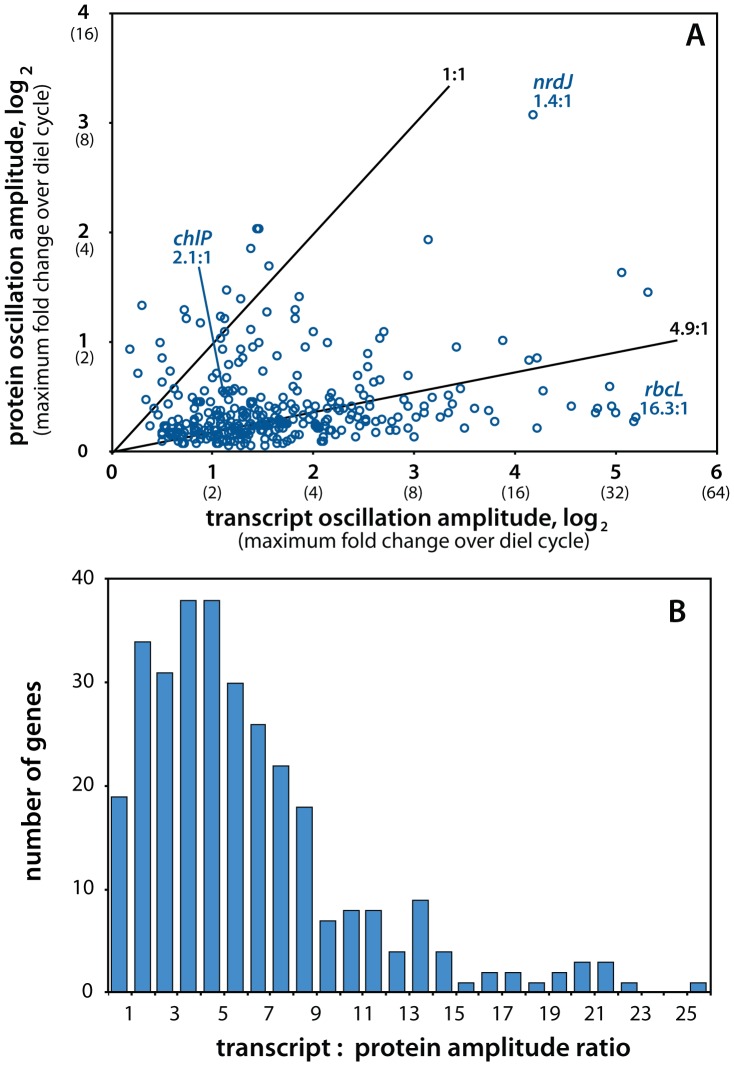
Methodology/principal findings: Using RNA-sequencing transcriptomics and mass spectrometry-based quantitative proteomics, we measured timecourses of paired mRNA-protein abundances for 312 genes every 2 hours over a light-dark cycle. These temporal expression patterns reveal strong oscillations in transcript abundance that are broadly damped at the protein level, with mRNA levels varying on average 2.3 times more than the corresponding protein. The single strongest observed protein-level oscillation is in a ribonucleotide reductase, which may reflect a defense strategy against phage infection. The peak in abundance of most proteins also lags that of their transcript by 2-8 hours, and the two are completely antiphase for some genes. While abundant antisense RNA was detected, it apparently does not account for the observed divergences between expression levels. The redirection of flux through central carbon metabolism from daytime carbon fixation to nighttime respiration is associated with quite small changes in relative enzyme abundances.
Conclusions/significance: Our results indicate that expression responses to periodic stimuli that are common in natural ecosystems (such as the diel cycle) can diverge significantly between the mRNA and protein levels. Protein expression patterns that are distinct from those of cognate mRNA have implications for the interpretation of transcriptome and metatranscriptome data in terms of cellular metabolism and its biogeochemical impact.
Conflict of interest statement
Figures




References
-
- Crick F (1970) Central dogma of molecular biology. Nature 227: 561–563. - PubMed
-
- Picard F, Dressaire C, Girbal L, Cocaign-Bousquet M (2009) Examination of post-transcriptional regulations in prokaryotes by integrative biology. Comptes rendus biologies 332: 958–973. - PubMed
-
- Evguenieva-Hackenberg E, Klug G (2011) New aspects of RNA processing in prokaryotes. Current Opinion in Microbiology 14: 587–592. - PubMed
-
- Lee PS, Shaw LB, Choe LH, Mehra A, Hatzimanikatis V, et al. (2003) Insights into the relation between mrna and protein expression patterns: II. Experimental observations in Escherichia coli . Biotechnology and bioengineering 84: 834–841. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
