

Potent adjuvanticity of a pure TLR7-agonistic imidazoquinoline dendrimer
- PMID: 22952720
- PMCID: PMC3429503
- DOI: 10.1371/journal.pone.0043612
Potent adjuvanticity of a pure TLR7-agonistic imidazoquinoline dendrimer
Abstract
Engagement of toll-like receptors (TLRs) serve to link innate immune responses with adaptive immunity and can be exploited as powerful vaccine adjuvants for eliciting both primary and anamnestic immune responses. TLR7 agonists are highly immunostimulatory without inducing dominant proinflammatory cytokine responses. We synthesized a dendrimeric molecule bearing six units of a potent TLR7/TLR8 dual-agonistic imidazoquinoline to explore if multimerization of TLR7/8 would result in altered activity profiles. A complete loss of TLR8-stimulatory activity with selective retention of the TLR7-agonistic activity was observed in the dendrimer. This was reflected by a complete absence of TLR8-driven proinflammatory cytokine and interferon (IFN)-γ induction in human PBMCs, with preservation of TLR7-driven IFN-α induction. The dendrimer was found to be superior to the imidazoquinoline monomer in inducing high titers of high-affinity antibodies to bovine α-lactalbumin. Additionally, epitope mapping experiments showed that the dendrimer induced immunoreactivity to more contiguous peptide epitopes along the amino acid sequence of the model antigen.
Conflict of interest statement
Figures
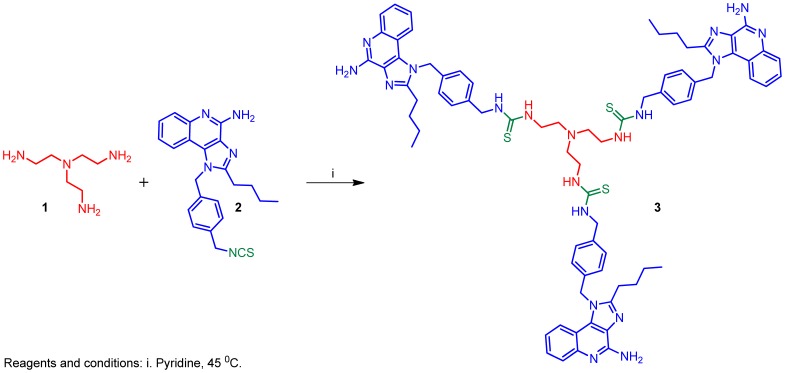

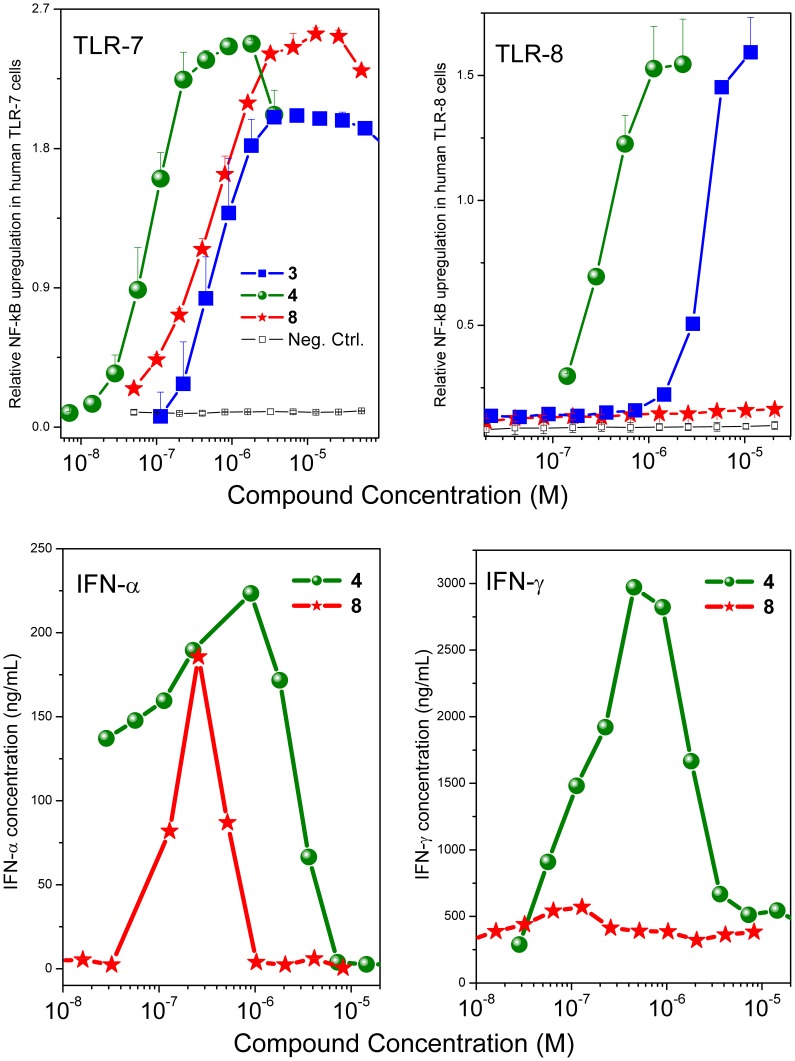





References
-
- Kawai T, Akira S (2007) TLR signaling. Semin Immunol 19: 24–32. - PubMed
-
- Kumagai Y, Takeuchi O, Akira S (2008) Pathogen recognition by innate receptors. J Infect Chemother 14: 86–92. - PubMed
-
- Akira S (1902) Toll-like receptors and innate immunity. Adv Immunol 78: 1–56. - PubMed
-
- Akira S, Takeda K, Kaisho T (2001) Toll-like receptors: critical proteins linking innate and acquired immunity. Nature Immunol 2: 675–680. - PubMed
-
- Cottalorda A, Verschelde C, Marcais A, Tomkowiak M, Musette P, et al. (2006) TLR2 engagement on CD8 T cells lowers the threshold for optimal antigen-induced T cell activation. Eur J Immunol 36: 1684–1693. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
