

Evolution of multiple additive loci caused divergence between Drosophila yakuba and D. santomea in wing rowing during male courtship
- PMID: 22952802
- PMCID: PMC3431401
- DOI: 10.1371/journal.pone.0043888
Evolution of multiple additive loci caused divergence between Drosophila yakuba and D. santomea in wing rowing during male courtship
Abstract
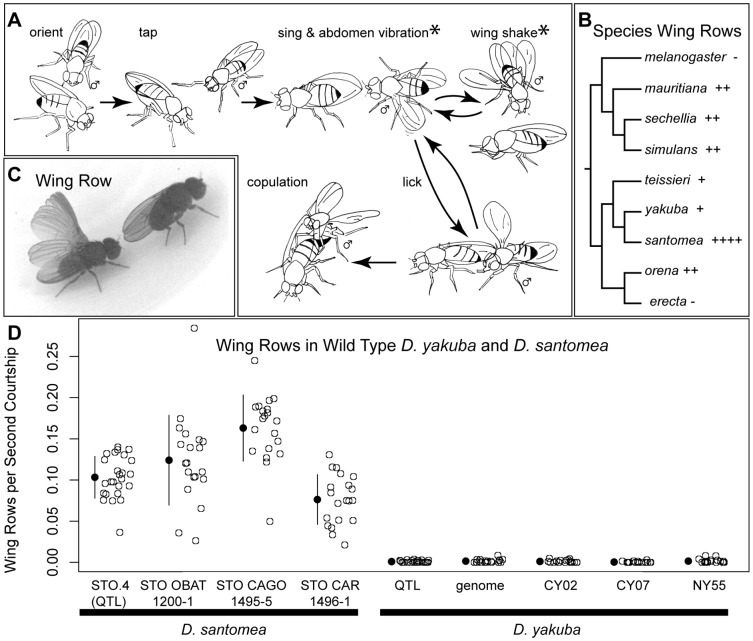
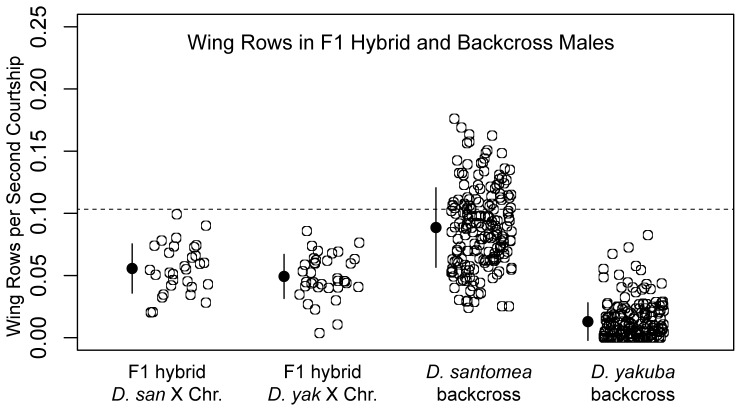
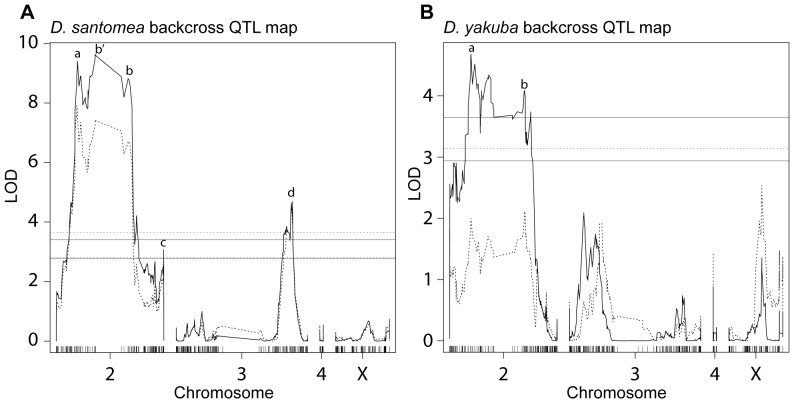
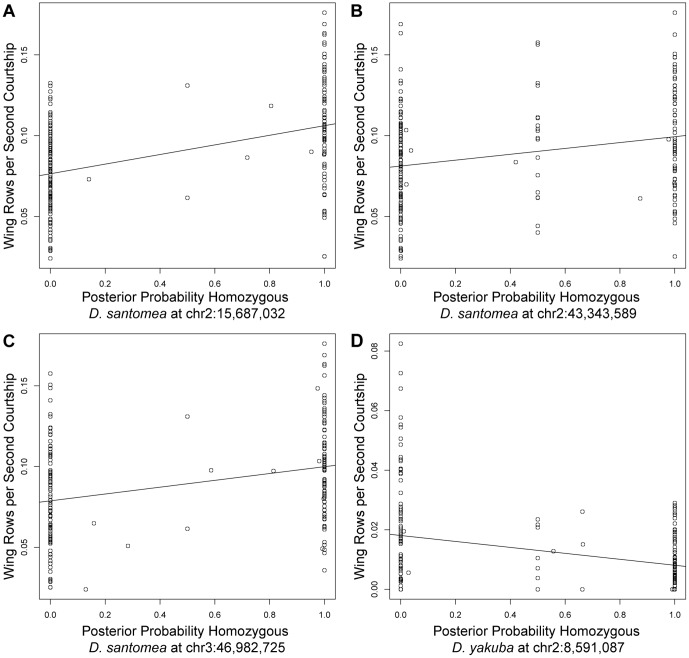
In Drosophila, male flies perform innate, stereotyped courtship behavior. This innate behavior evolves rapidly between fly species, and is likely to have contributed to reproductive isolation and species divergence. We currently understand little about the neurobiological and genetic mechanisms that contributed to the evolution of courtship behavior. Here we describe a novel behavioral difference between the two closely related species D. yakuba and D. santomea: the frequency of wing rowing during courtship. During courtship, D. santomea males repeatedly rotate their wing blades to face forward and then back (rowing), while D. yakuba males rarely row their wings. We found little intraspecific variation in the frequency of wing rowing for both species. We exploited multiplexed shotgun genotyping (MSG) to genotype two backcross populations with a single lane of Illumina sequencing. We performed quantitative trait locus (QTL) mapping using the ancestry information estimated by MSG and found that the species difference in wing rowing mapped to four or five genetically separable regions. We found no evidence that these loci display epistasis. The identified loci all act in the same direction and can account for most of the species difference.
Conflict of interest statement
Figures
References
-
- West-Eberhard MJ (1983) Sexual selection, social competition, and speciation. Q Rev Biol 58: 155–183.
-
- Martins EP, editor (1996) Phylogenies and the Comparative Method in Animal Behavior. New York: Oxford University Press, Inc. 432p.
-
- Williams TM, Carroll SB (2009) Genetic and molecular insights into the development and evolution of sexual dimorphism. Nat Rev Genet 10: 797–804. - PubMed
-
- Shubin N, Tabin C, Carroll S (2009) Deep homology and the origins of evolutionary novelty. Nature 457: 818–823. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
