

Age-dependent changes in innate immune phenotype and function in rhesus macaques (Macaca mulatta)
- PMID: 22953039
- PMCID: PMC3417700
- DOI: 10.3402/pba.v2i0.18052
Age-dependent changes in innate immune phenotype and function in rhesus macaques (Macaca mulatta)
Abstract
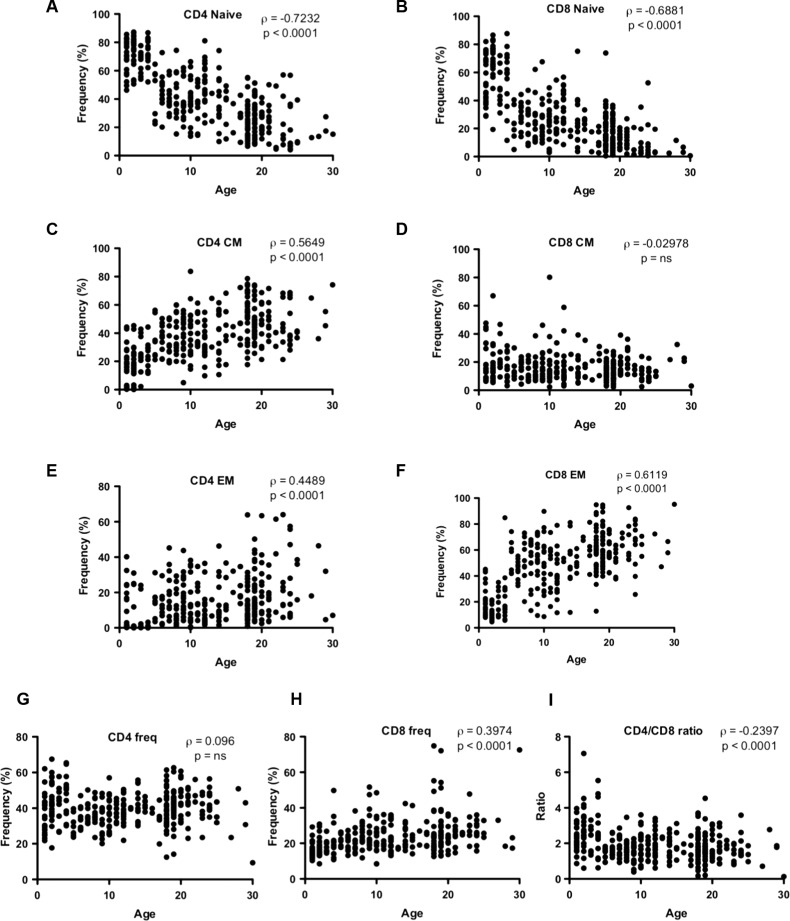
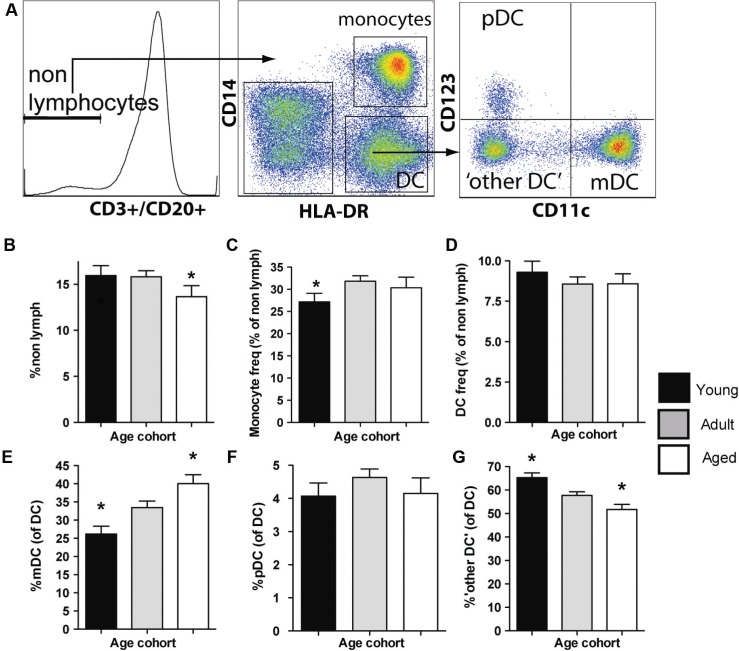
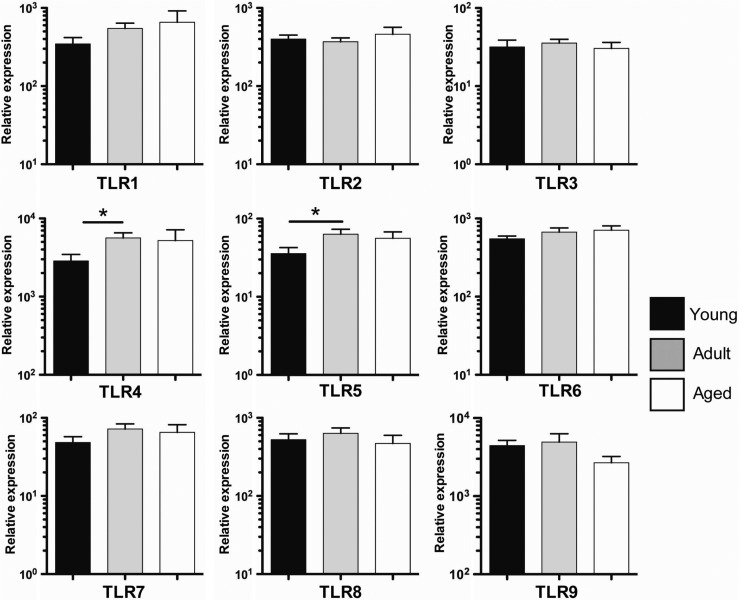
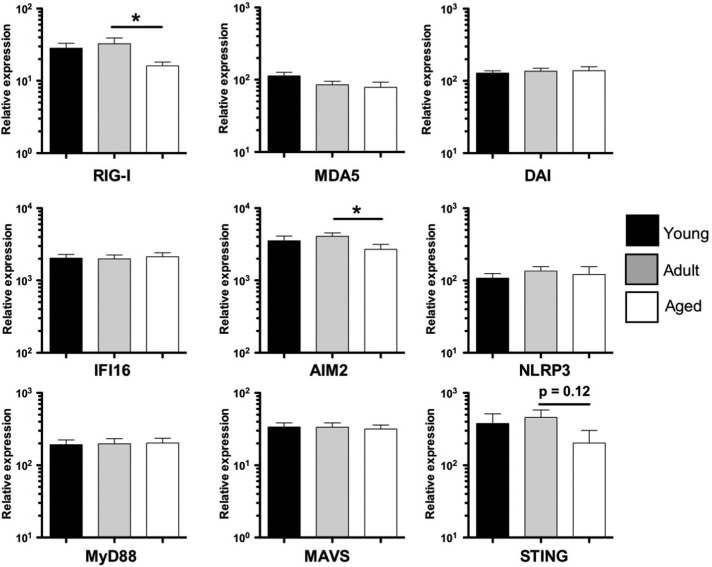
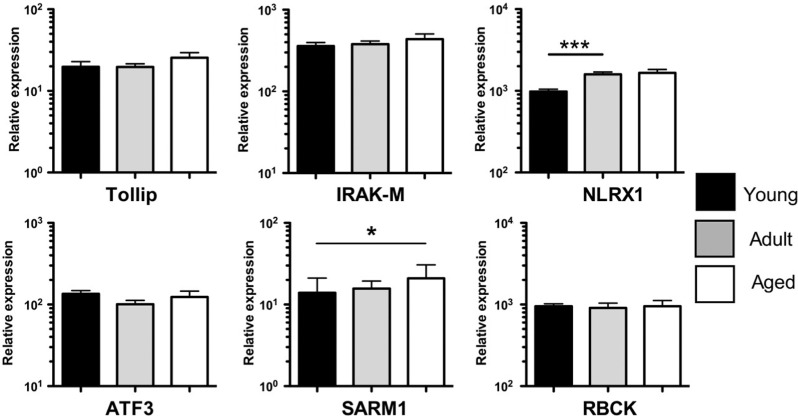
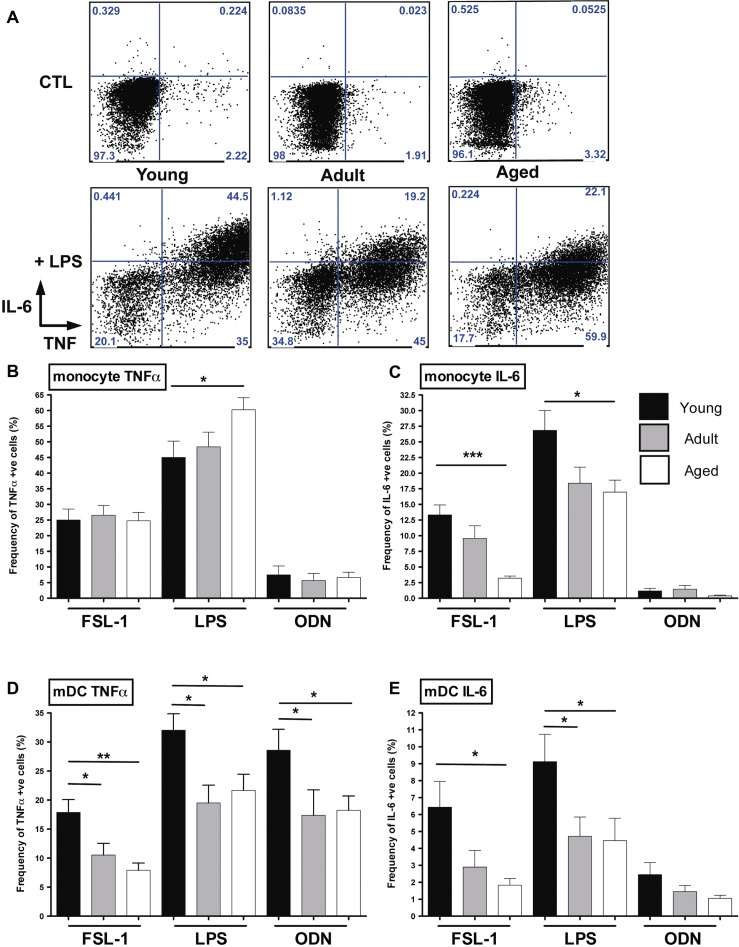
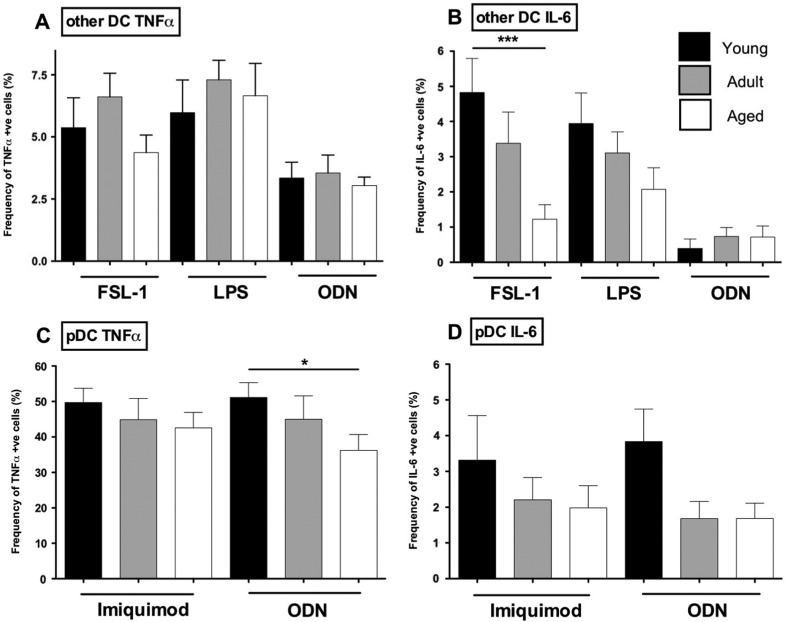
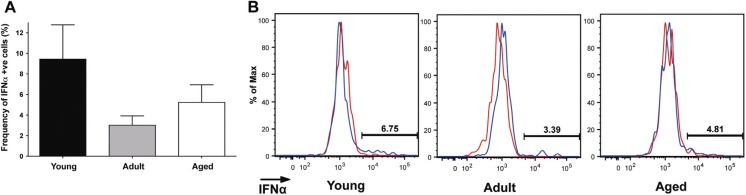
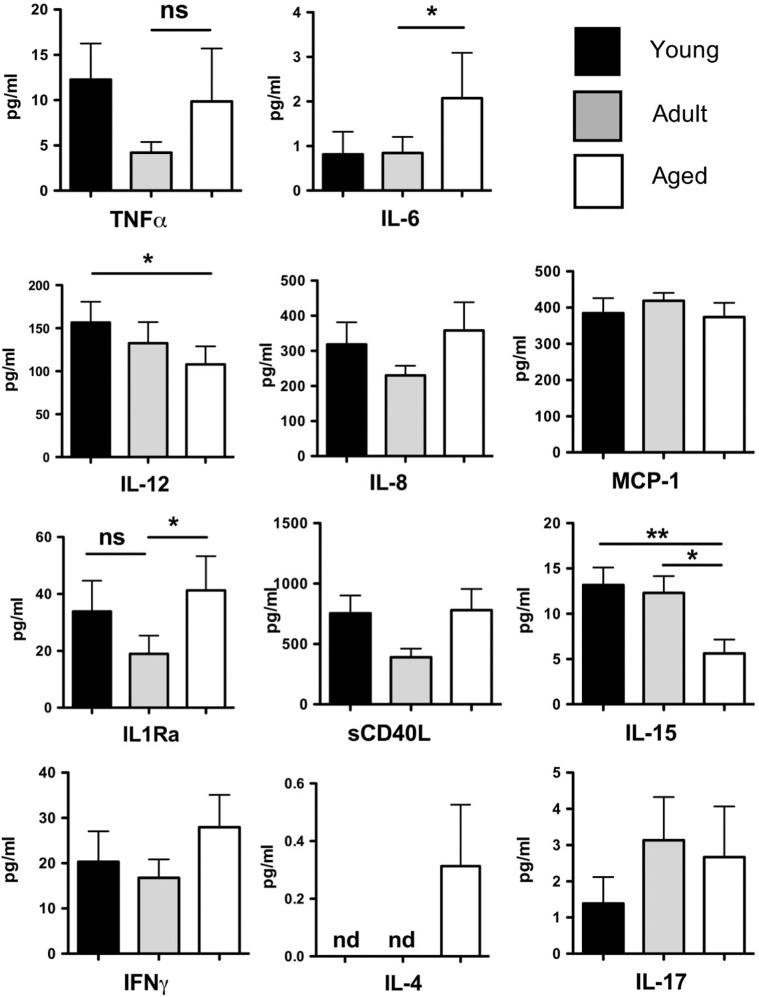
Aged individuals are more susceptible to infections due to a general decline in immune function broadly referred to as immune senescence. While age-related changes in the adaptive immune system are well documented, aging of the innate immune system remains less well understood, particularly in nonhuman primates. A more robust understanding of age-related changes in innate immune function would provide mechanistic insight into the increased susceptibility of the elderly to infection. Rhesus macaques have proved a critical translational model for aging research, and present a unique opportunity to dissect age-dependent modulation of the innate immune system. We examined age-related changes in: (i) innate immune cell frequencies; (ii) expression of pattern recognition receptors (PRRs) and innate signaling molecules; (iii) cytokine responses of monocytes and dendritic cells (DC) following stimulation with PRR agonists; and (iv) plasma cytokine levels in this model. We found marked changes in both the phenotype and function of innate immune cells. This included an age-associated increased frequency of myeloid DC (mDC). Moreover, we found toll-like receptor (TLR) agonists lipopolysaccharide (TLR4), fibroblast stimulating ligand-1 (TLR2/6), and ODN2006 (TLR7/9) induced reduced cytokine responses in aged mDC. Interestingly, with the exception of the monocyte-derived TNFα response to LPS, which increased with age, TNFα, IL-6, and IFNα responses declined with age. We also found that TLR4, TLR5, and innate negative regulator, sterile alpha and TIR motif containing protein (SARM), were all expressed at lower levels in young animals. By contrast, absent in melanoma 2 and retinoic acid-inducible gene I expression was lowest in aged animals. Together, these observations indicate that several parameters of innate immunity are significantly modulated by age and contribute to differential immune function in aged macaques.
Keywords: immune senscence; innate immunity; myeloid; nonhuman primate; pattern recognition receptor.
Figures
References
-
- Siegrist CA, Aspinall R. B-cell responses to vaccination at the extremes of age. Nat Rev Immunol. 2009;9:185–94. - PubMed
-
- Siegrist CA. Neonatal and early life vaccinology. Vaccine. 2001;19:3331–46. - PubMed
-
- Adkins B, Leclerc C, Marshall-Clarke S. Neonatal adaptive immunity comes of age. Nat Rev Immunol. 2004;4:553–64. - PubMed
-
- Yoshikawa TT. Epidemiology and unique aspects of aging and infectious diseases. Clin Infect Dis. 2000;30:931–33. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources