

Decreased mitochondrial apoptotic priming underlies stroma-mediated treatment resistance in chronic lymphocytic leukemia
- PMID: 22955911
- PMCID: PMC3482860
- DOI: 10.1182/blood-2012-02-414060
Decreased mitochondrial apoptotic priming underlies stroma-mediated treatment resistance in chronic lymphocytic leukemia
Abstract
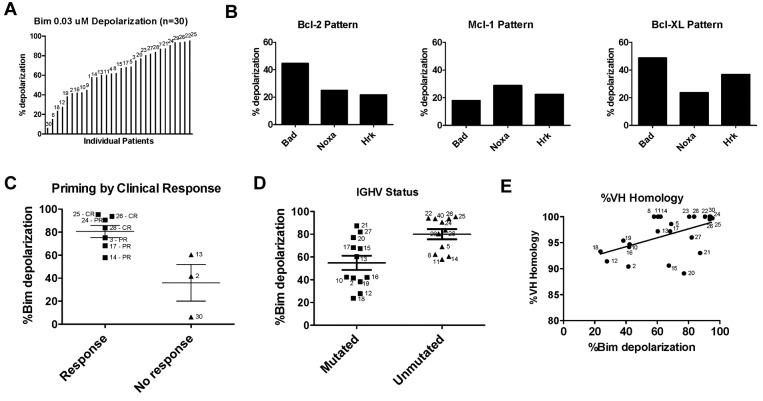
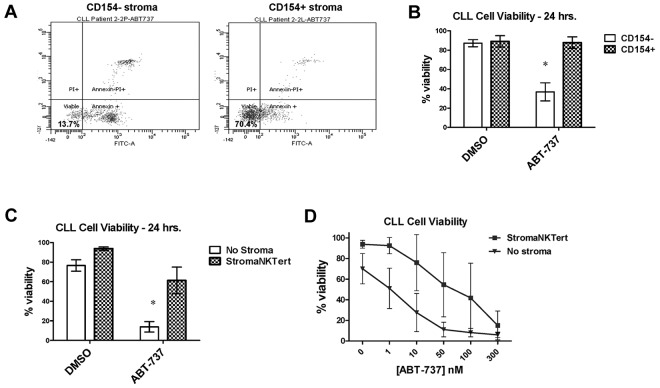
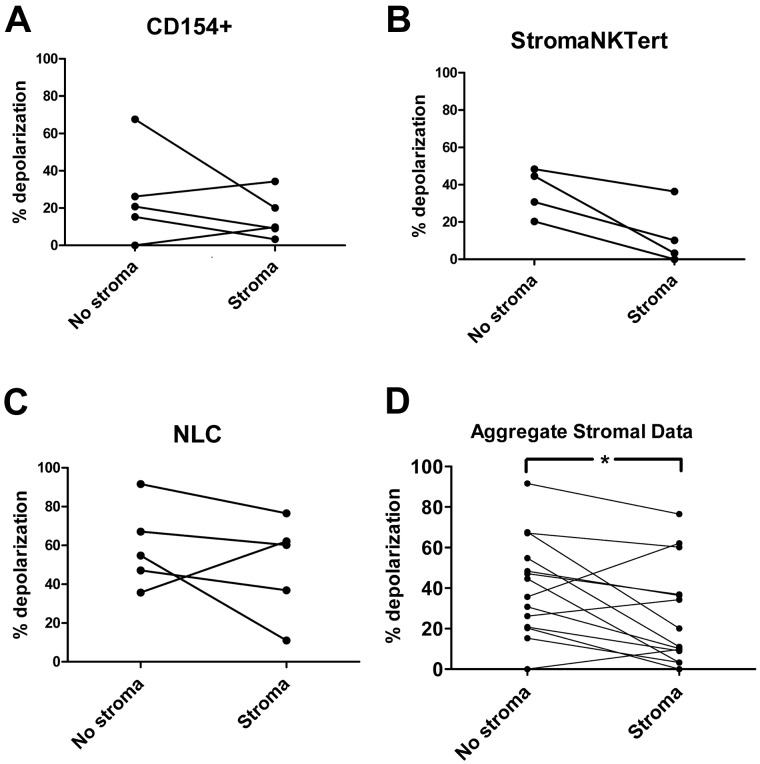
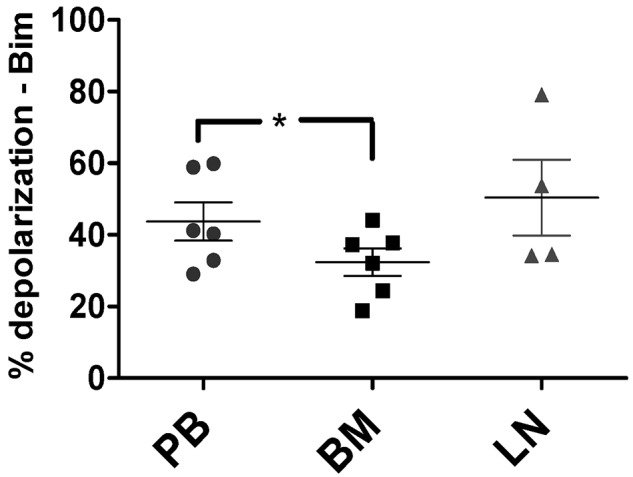
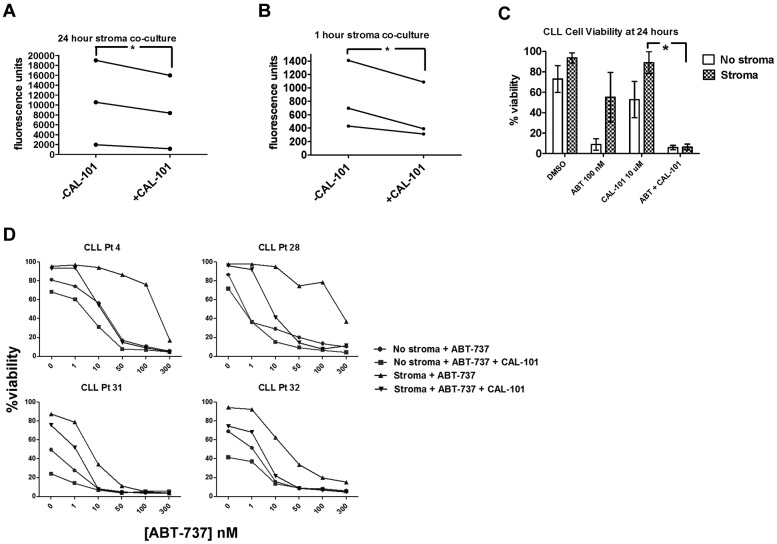
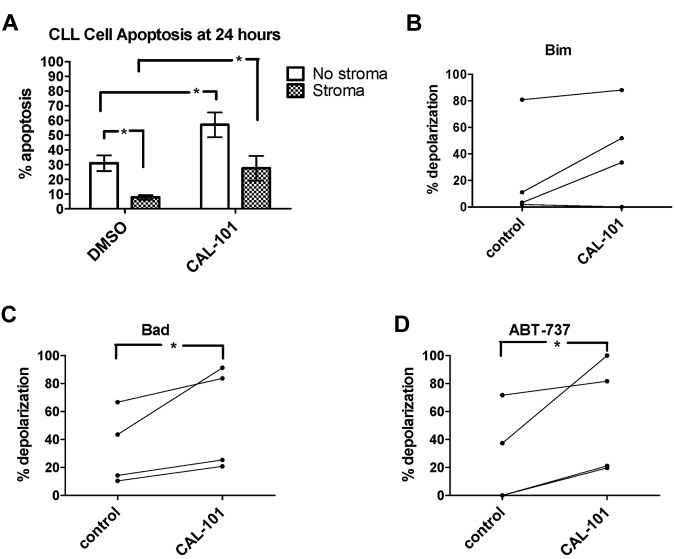
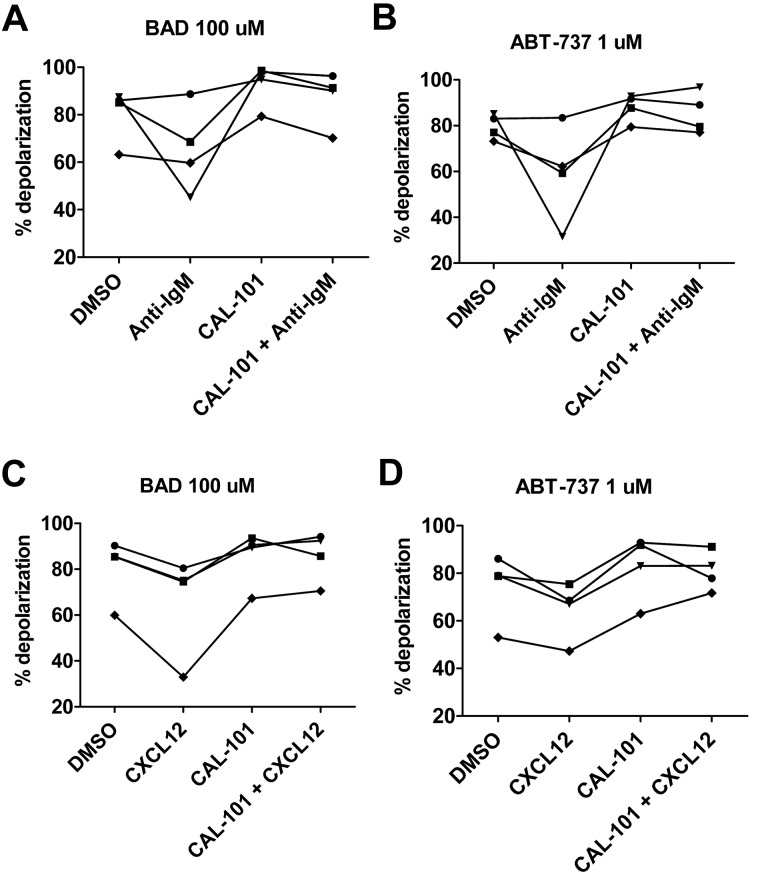
Stroma induces treatment resistance in chronic lymphocytic leukemia (CLL), possibly because of alterations in the BCL-2 family of proteins, which are key regulators of apoptosis. We previously developed BH3 profiling, a functional assay that assesses mitochondrial depolarization in response to BH3-only peptides, to measure "apoptotic priming," the proximity of a cell to the apoptotic threshold. In the present study, we use BH3 profiling to show that CLL cells from the PB are highly primed. Increased priming is associated with improved clinical response and, unexpectedly, with unmutated IGHV status. Coculturing CLL cells in vitro with stroma decreases priming. Using matched PB, BM, and lymph node compartment samples, we found in vivo that BM-derived CLL cells are the least primed. CLL cells cocultured with stroma were treated with the PI3K δ-isoform inhibitor CAL-101 (GS1101). CAL-101 caused CLL cell de-adhesion, leading to increased CLL cell priming. Stimulation of CLL cells with anti-IgM or CXCL12 caused decreased priming that could be reversed by CAL-101. Our results show that inhibition of stromal interactions leading to displacement of CLL cells into the blood by CAL-101 in vivo may increase CLL cell priming, suggesting a mechanism by which agents inducing lymphocyte redistribution might facilitate improved clinical response when used in combination with other therapies.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
