

BICD2, dynactin, and LIS1 cooperate in regulating dynein recruitment to cellular structures
- PMID: 22956769
- PMCID: PMC3484101
- DOI: 10.1091/mbc.E12-03-0210
BICD2, dynactin, and LIS1 cooperate in regulating dynein recruitment to cellular structures
Abstract
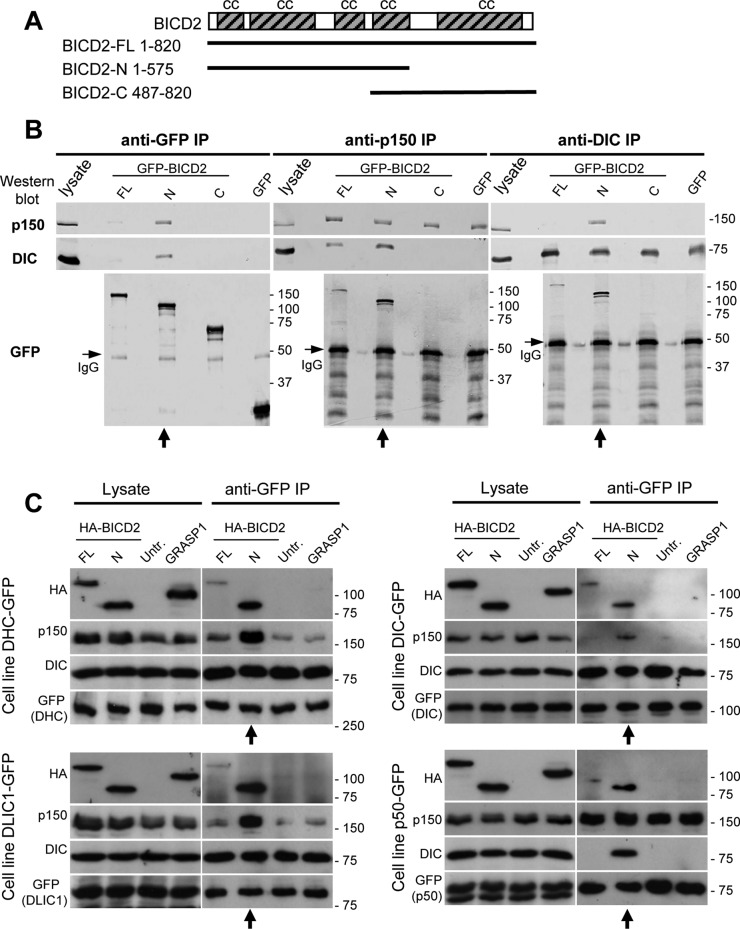
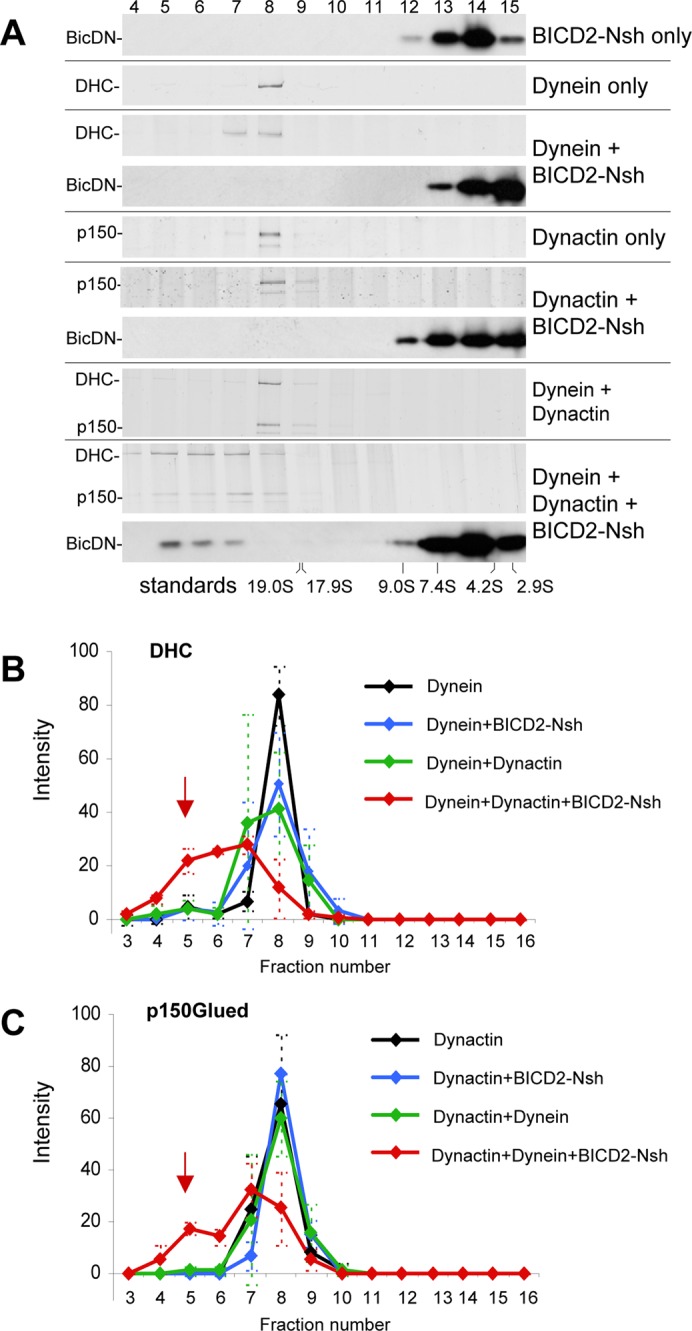
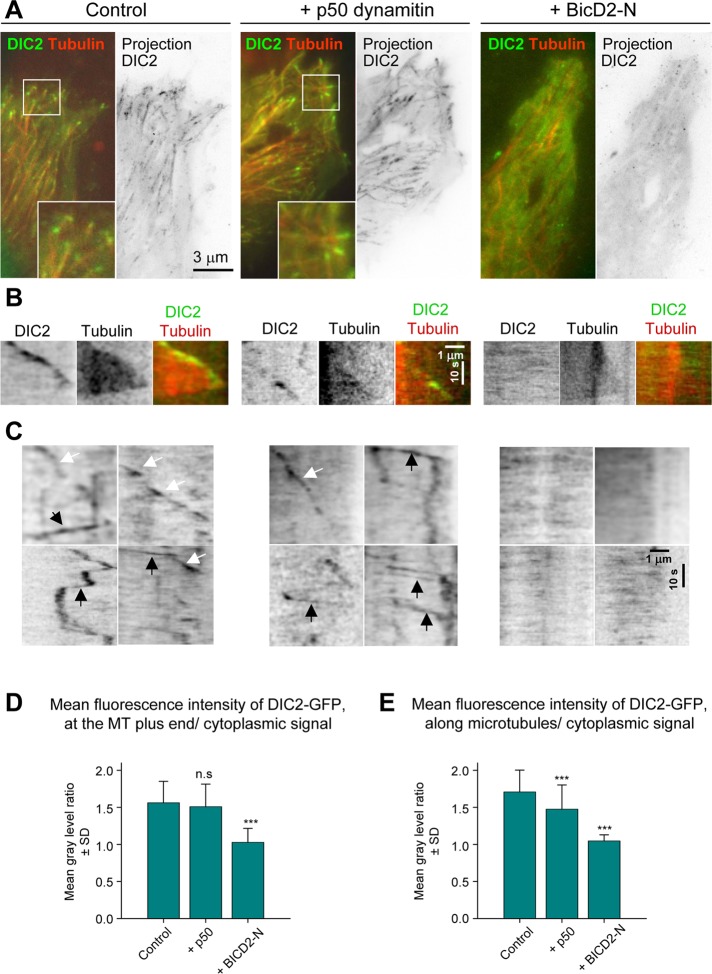
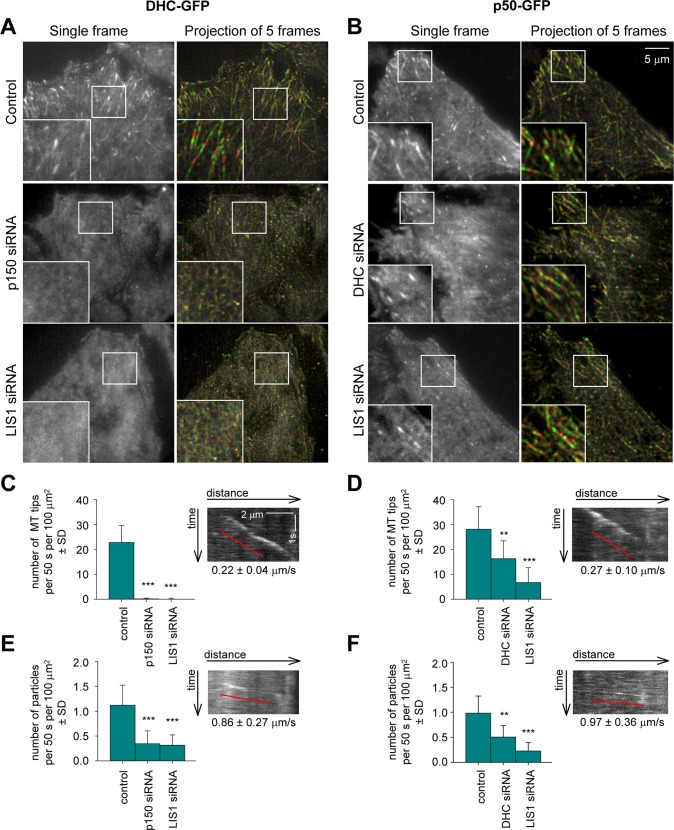
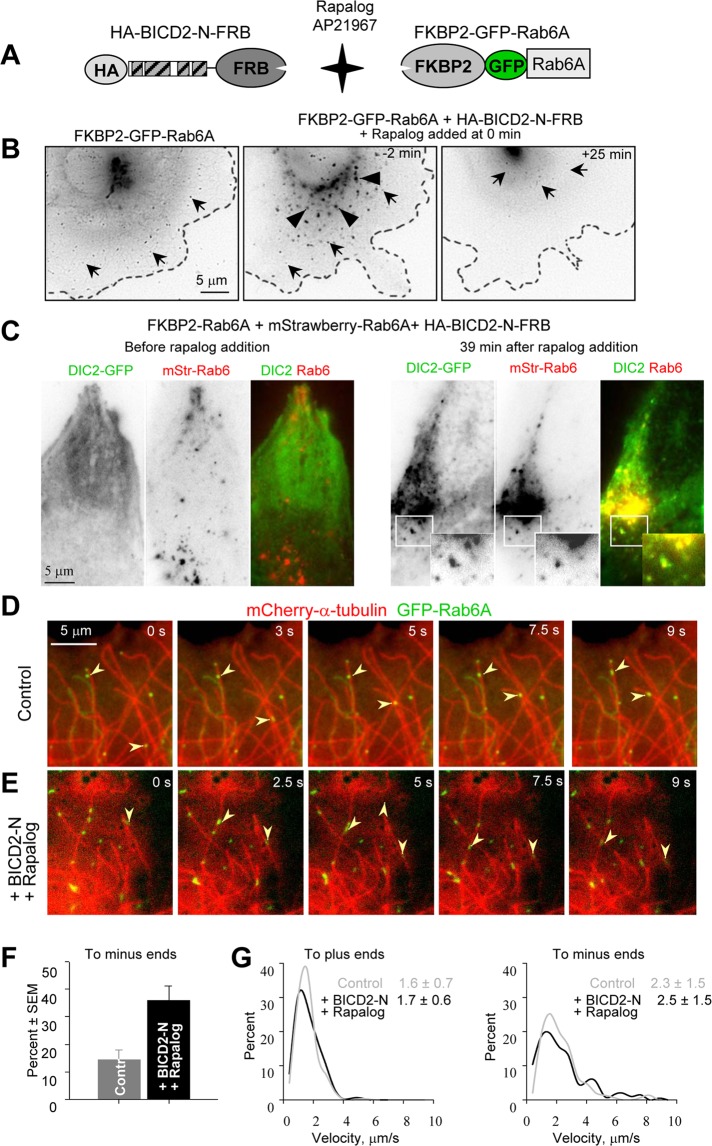
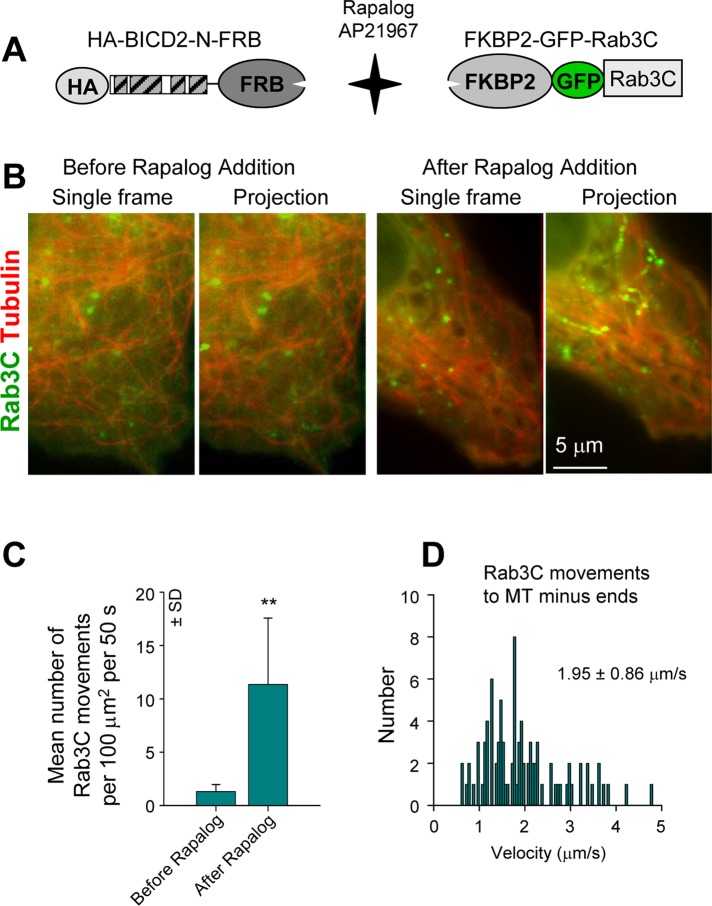
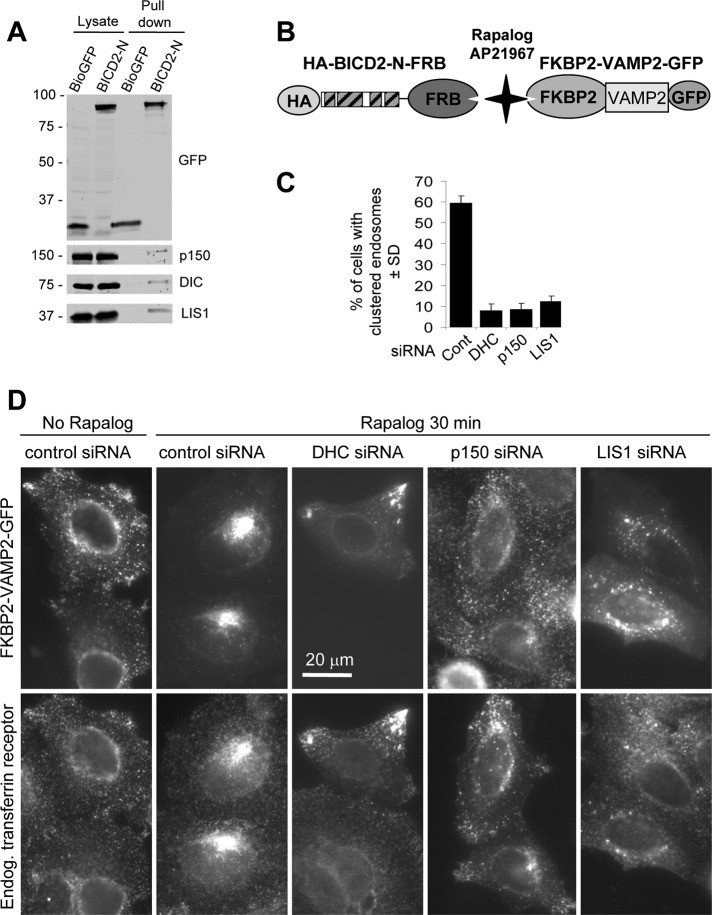
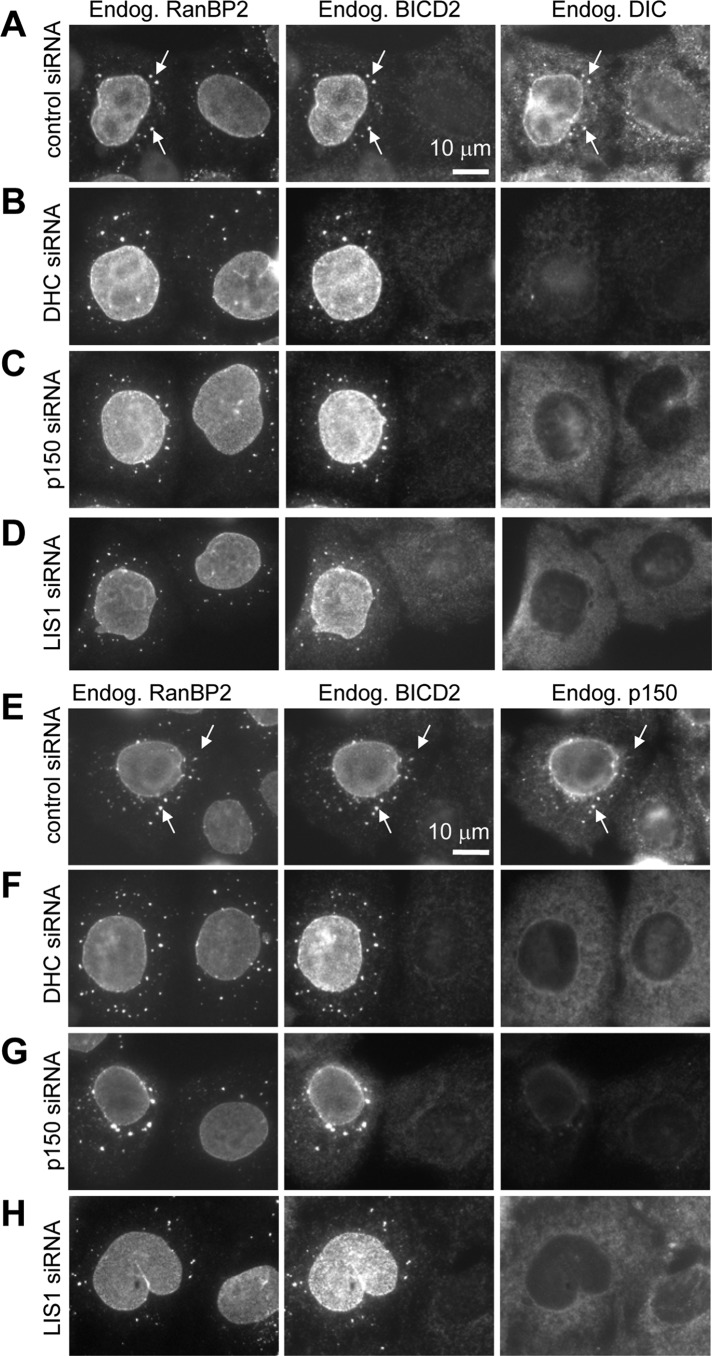
Cytoplasmic dynein is the major microtubule minus-end-directed cellular motor. Most dynein activities require dynactin, but the mechanisms regulating cargo-dependent dynein-dynactin interaction are poorly understood. In this study, we focus on dynein-dynactin recruitment to cargo by the conserved motor adaptor Bicaudal D2 (BICD2). We show that dynein and dynactin depend on each other for BICD2-mediated targeting to cargo and that BICD2 N-terminus (BICD2-N) strongly promotes stable interaction between dynein and dynactin both in vitro and in vivo. Direct visualization of dynein in live cells indicates that by itself the triple BICD2-N-dynein-dynactin complex is unable to interact with either cargo or microtubules. However, tethering of BICD2-N to different membranes promotes their microtubule minus-end-directed motility. We further show that LIS1 is required for dynein-mediated transport induced by membrane tethering of BICD2-N and that LIS1 contributes to dynein accumulation at microtubule plus ends and BICD2-positive cellular structures. Our results demonstrate that dynein recruitment to cargo requires concerted action of multiple dynein cofactors.
Figures
Comment in
-
Sorting out microtubule-based transport.Nat Rev Mol Cell Biol. 2021 Feb;22(2):73. doi: 10.1038/s41580-020-00320-y. Nat Rev Mol Cell Biol. 2021. PMID: 33288890 No abstract available.
References
-
- Bingham JB, King SJ, Schroer TA. Purification of dynactin and dynein from brain tissue. Methods Enzymol. 1998;298:171–184. - PubMed
-
- Bullock SL, Nicol A, Gross SP, Zicha D. Guidance of bidirectional motor complexes by mRNA cargoes through control of dynein number and activity. Curr Biol. 2006;16:1447–1452. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
