

Sox21 promotes hippocampal adult neurogenesis via the transcriptional repression of the Hes5 gene
- PMID: 22956844
- PMCID: PMC6621257
- DOI: 10.1523/JNEUROSCI.5803-11.2012
Sox21 promotes hippocampal adult neurogenesis via the transcriptional repression of the Hes5 gene
Abstract
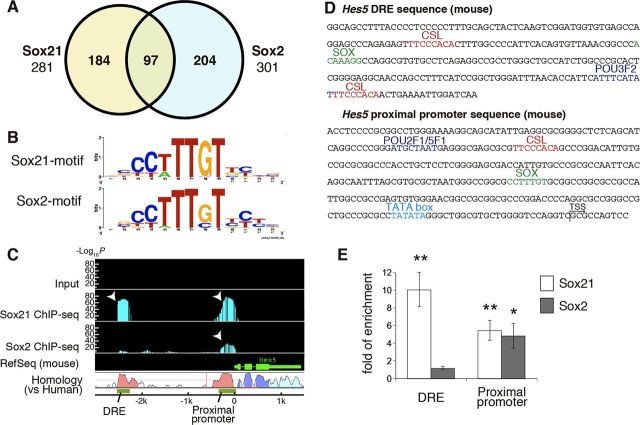
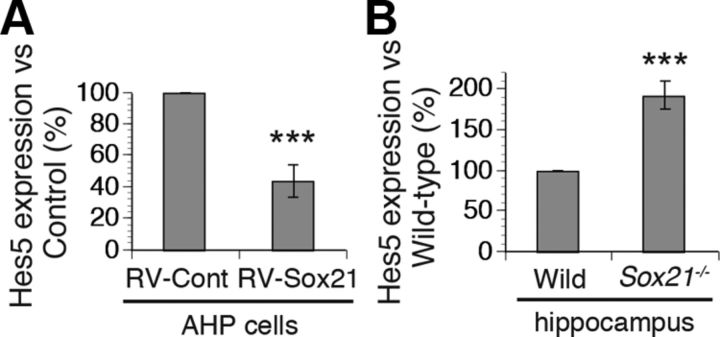
Despite the importance of the production of new neurons in the adult hippocampus, the transcription network governing this process remains poorly understood. The High Mobility Group (HMG)-box transcription factor, Sox2, and the cell surface activated transcriptional regulator, Notch, play important roles in CNS stem cells. Here, we demonstrate that another member of the SoxB (Sox1/Sox2/Sox3) transcription factor family, Sox21, is also a critical regulator of adult neurogenesis in mouse hippocampus. Loss of Sox21 impaired transition of progenitor cells from type 2a to type 2b, thereby reducing subsequent production of new neurons in the adult dentate gyrus. Analysis of the Sox21 binding sites in neural stem/progenitor cells indicated that the Notch-responsive gene, Hes5, was a target of Sox21. Sox21 repressed Hes5 gene expression at the transcriptional level. Simultaneous overexpression of Hes5 and Sox21 revealed that Hes5 was a downstream effector of Sox21 at the point where the Notch and Sox pathways intersect to control the number of neurons in the adult hippocampus. Therefore, Sox21 controls hippocampal adult neurogenesis via transcriptional repression of the Hes5 gene.
Figures
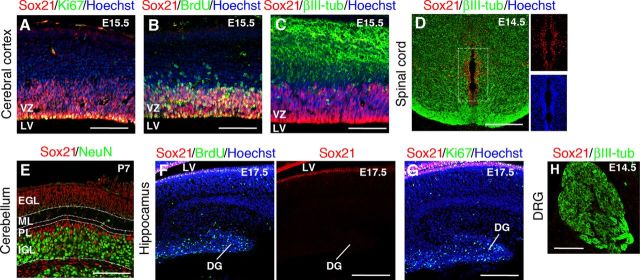

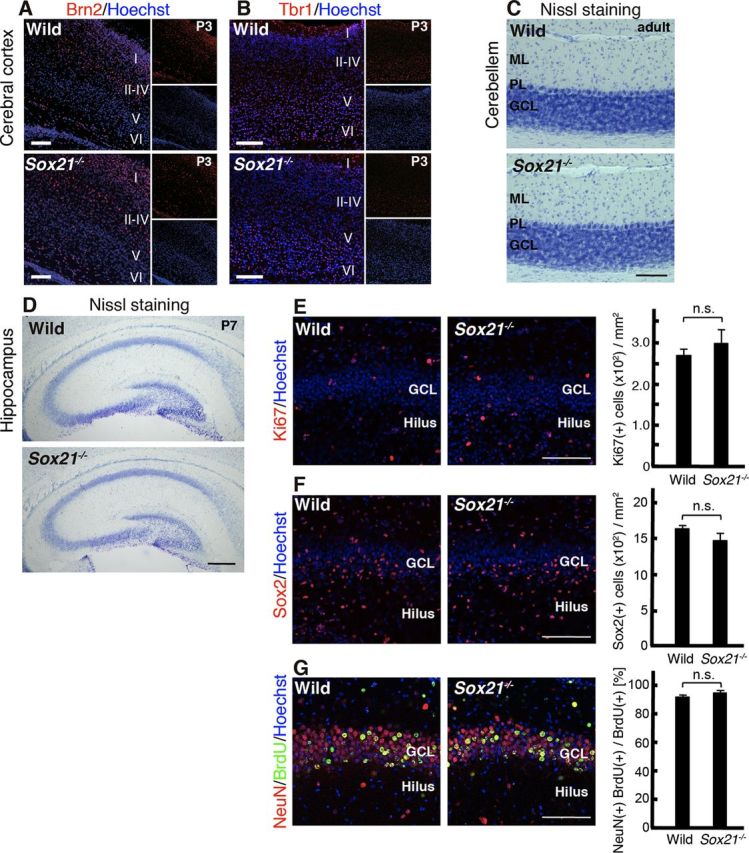
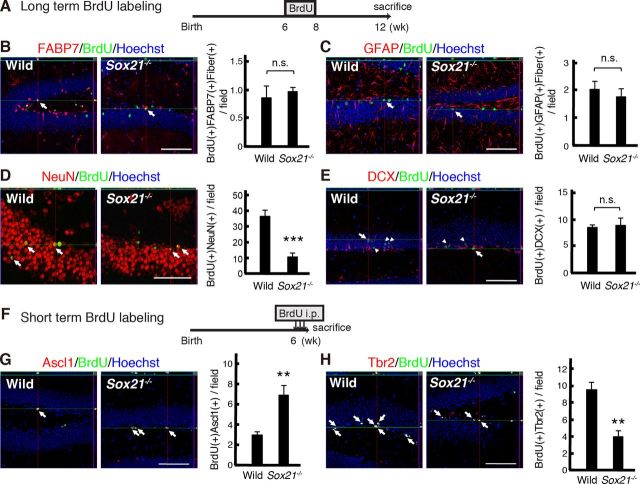

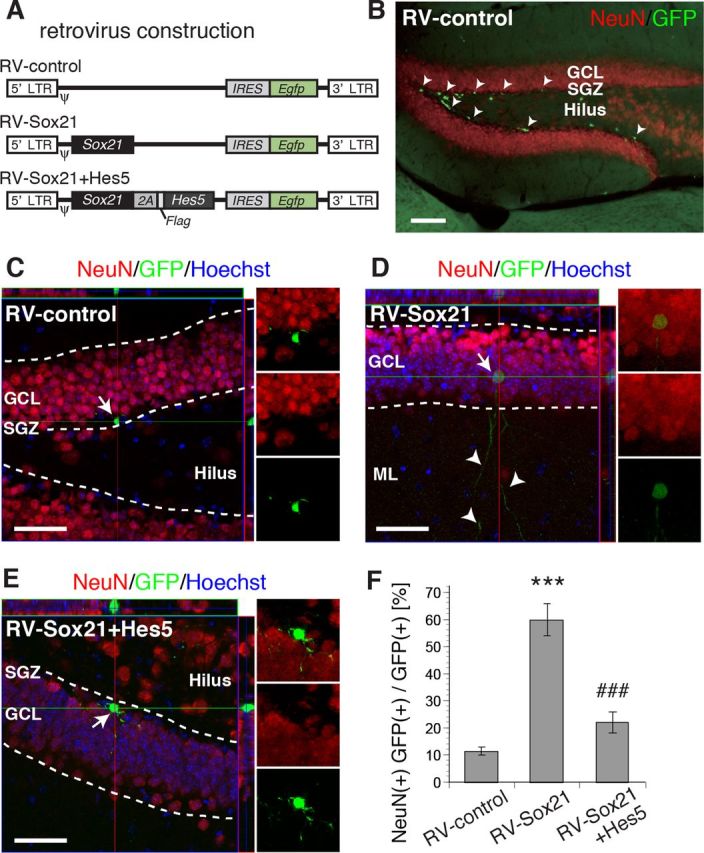



References
-
- Akazawa C, Sasai Y, Nakanishi S, Kageyama R. Molecular characterization of a rat negative regulator with a basic helix-loop-helix structure predominantly expressed in the developing nervous system. J Biol Chem. 1992;267:21879–21885. - PubMed
-
- Bailey TL, Elkan C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol. 1994;2:28–36. - PubMed
-
- Bani-Yaghoub M, Tremblay RG, Lei JX, Zhang D, Zurakowski B, Sandhu JK, Smith B, Ribecco-Lutkiewicz M, Kennedy J, Walker PR, Sikorska M. Role of Sox2 in the development of the mouse neocortex. Dev Biol. 2006;295:52–66. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous