

CPSF6 defines a conserved capsid interface that modulates HIV-1 replication
- PMID: 22956906
- PMCID: PMC3431306
- DOI: 10.1371/journal.ppat.1002896
CPSF6 defines a conserved capsid interface that modulates HIV-1 replication
Abstract
The HIV-1 genome enters cells inside a shell comprised of capsid (CA) protein. Variation in CA sequence alters HIV-1 infectivity and escape from host restriction factors. However, apart from the Cyclophilin A-binding loop, CA has no known interfaces with which to interact with cellular cofactors. Here we describe a novel protein-protein interface in the N-terminal domain of HIV-1 CA, determined by X-ray crystallography, which mediates both viral restriction and host cofactor dependence. The interface is highly conserved across lentiviruses and is accessible in the context of a hexameric lattice. Mutation of the interface prevents binding to and restriction by CPSF6-358, a truncated cytosolic form of the RNA processing factor, cleavage and polyadenylation specific factor 6 (CPSF6). Furthermore, mutations that prevent CPSF6 binding also relieve dependence on nuclear entry cofactors TNPO3 and RanBP2. These results suggest that the HIV-1 capsid mediates direct host cofactor interactions to facilitate viral infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
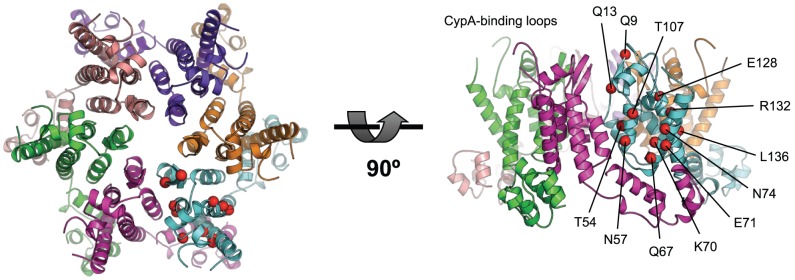
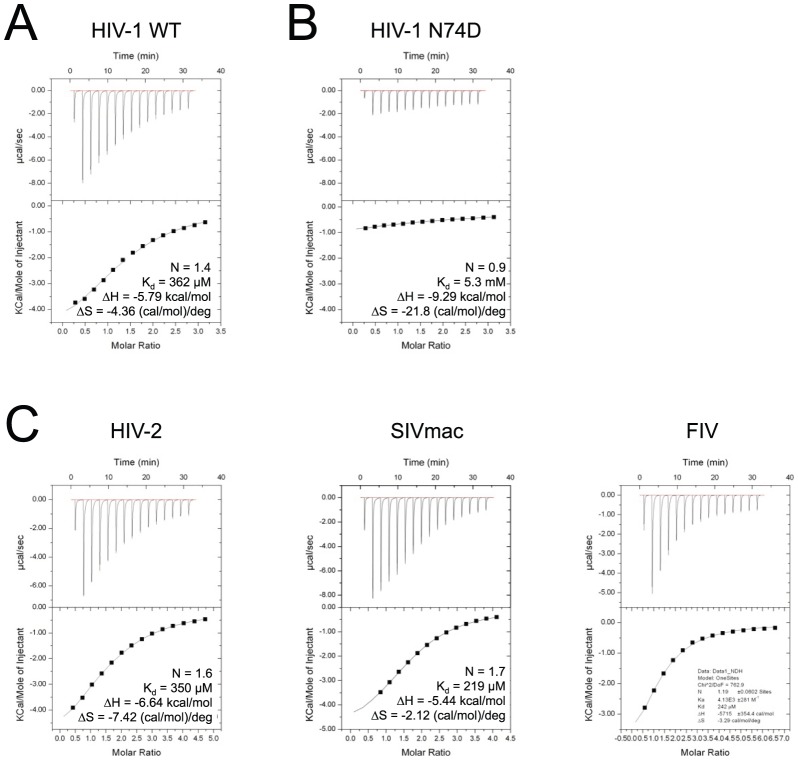
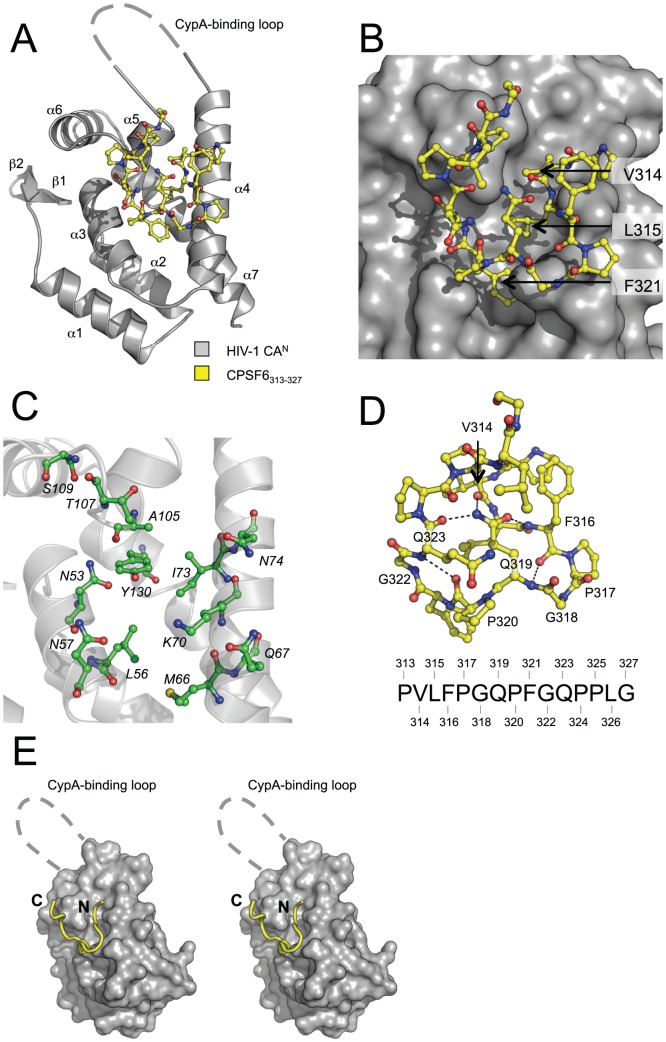
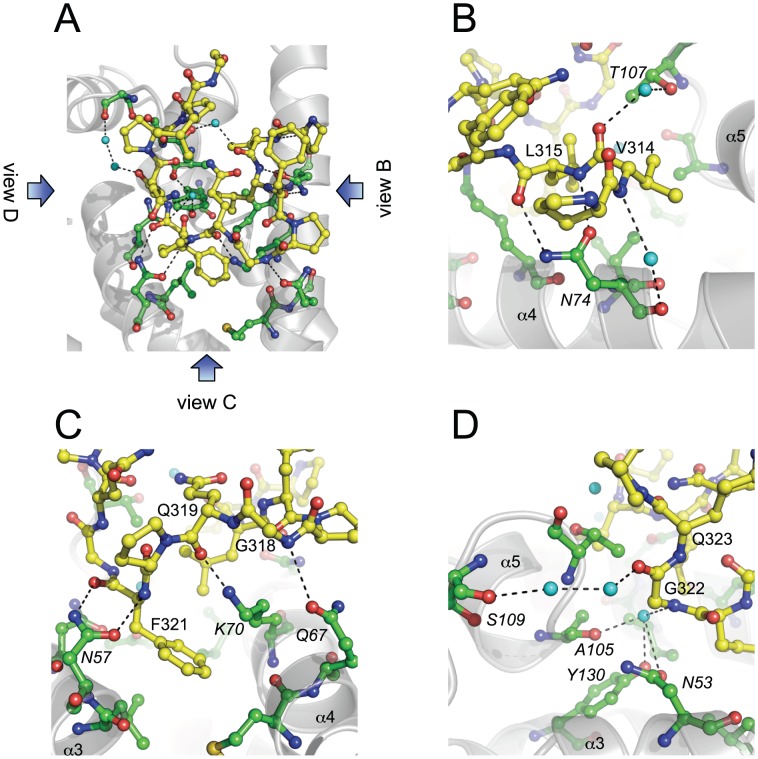
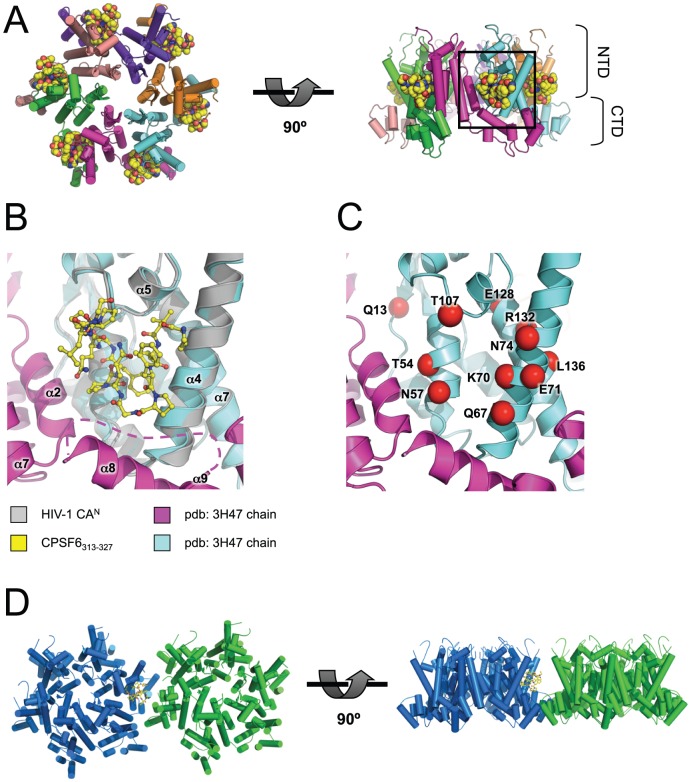
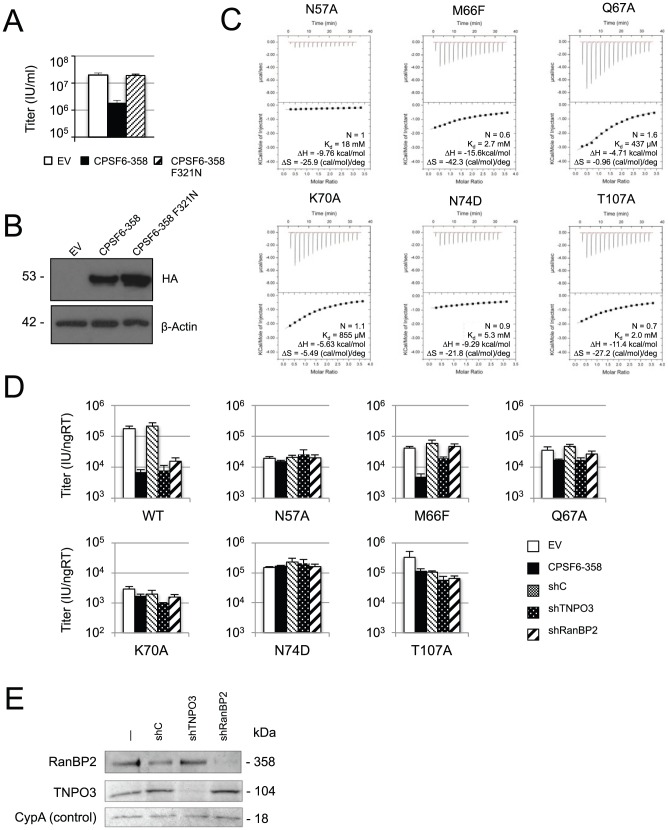
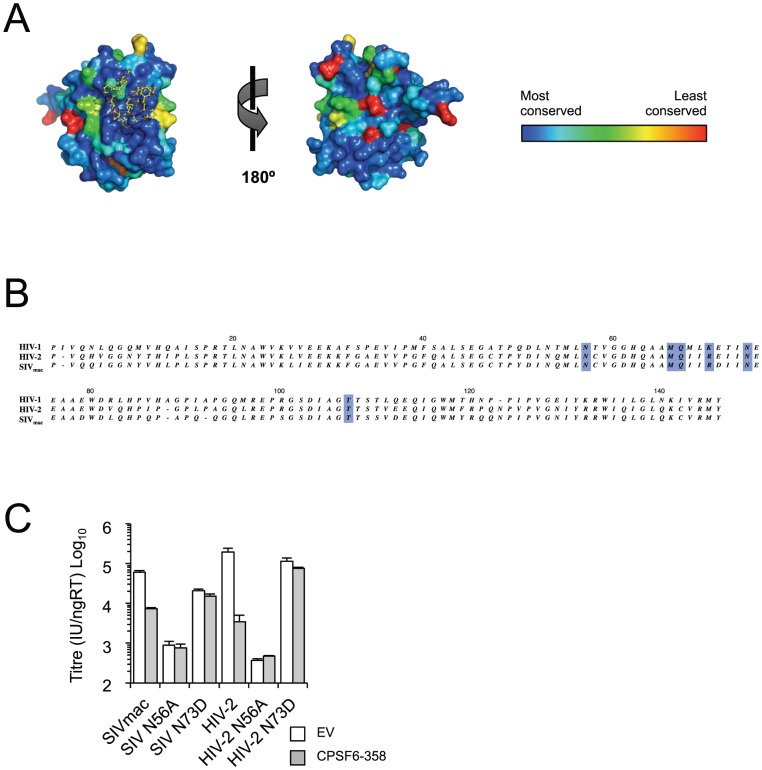
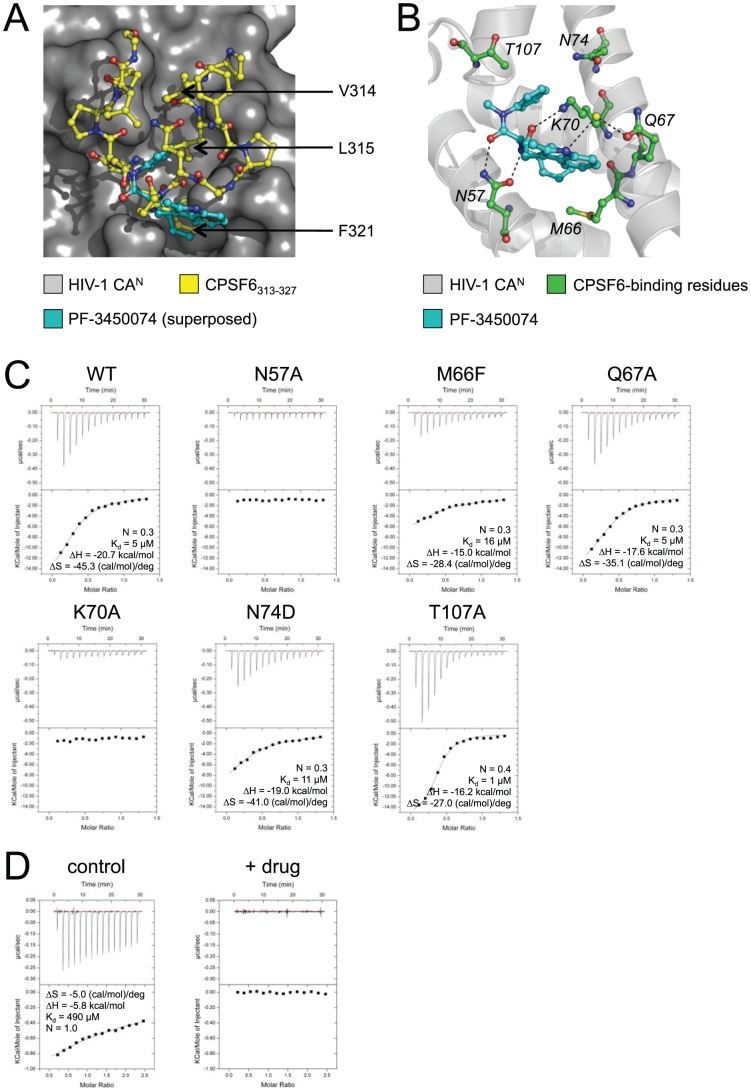
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
