

Drosophila melanogaster Acetyl-CoA-carboxylase sustains a fatty acid-dependent remote signal to waterproof the respiratory system
- PMID: 22956916
- PMCID: PMC3431307
- DOI: 10.1371/journal.pgen.1002925
Drosophila melanogaster Acetyl-CoA-carboxylase sustains a fatty acid-dependent remote signal to waterproof the respiratory system
Abstract
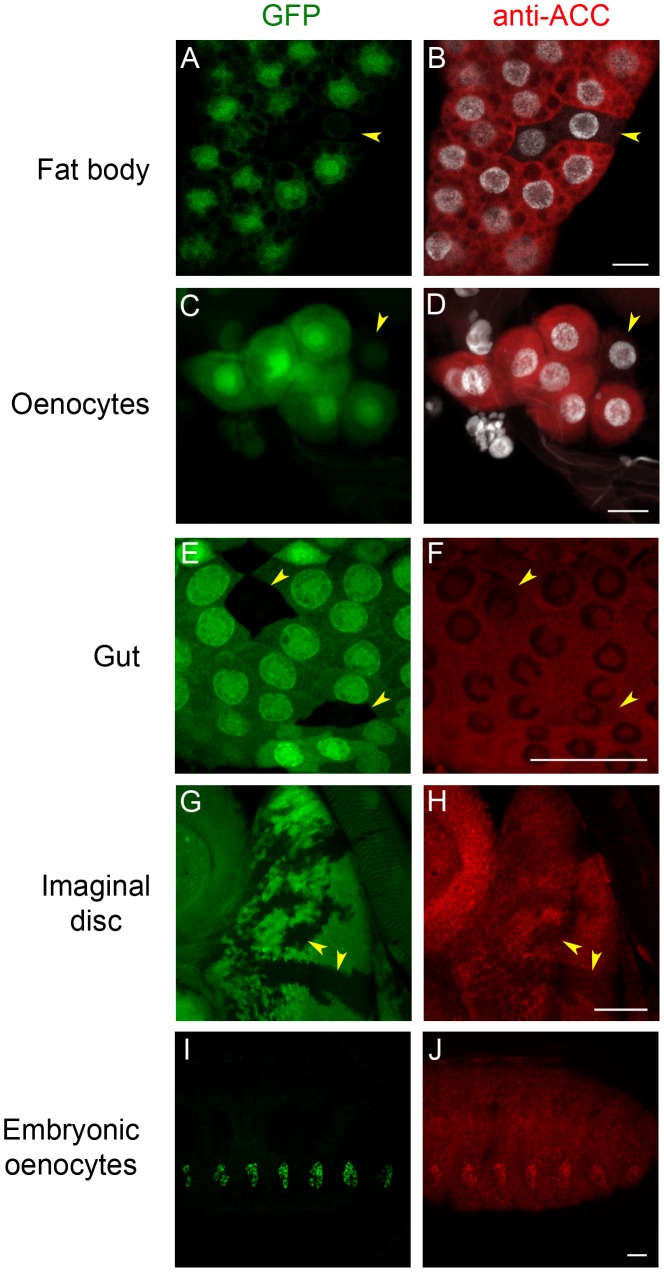
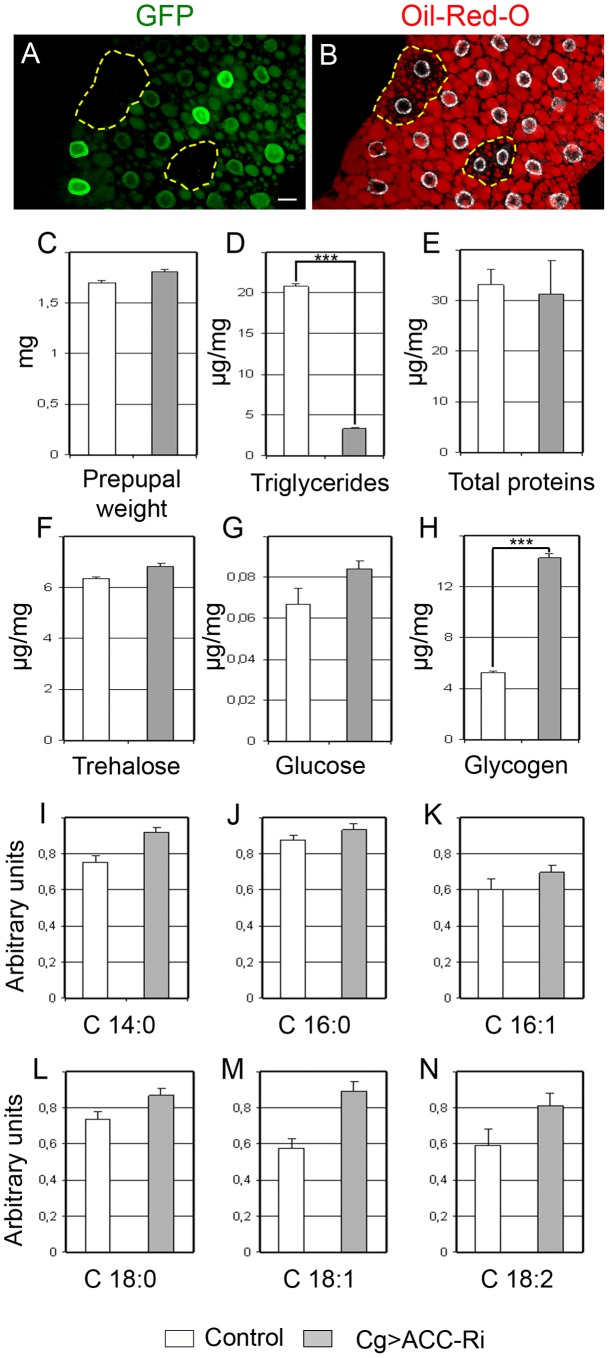
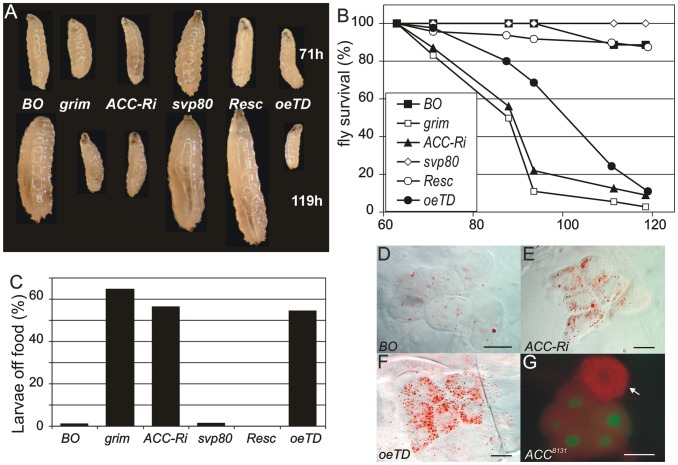
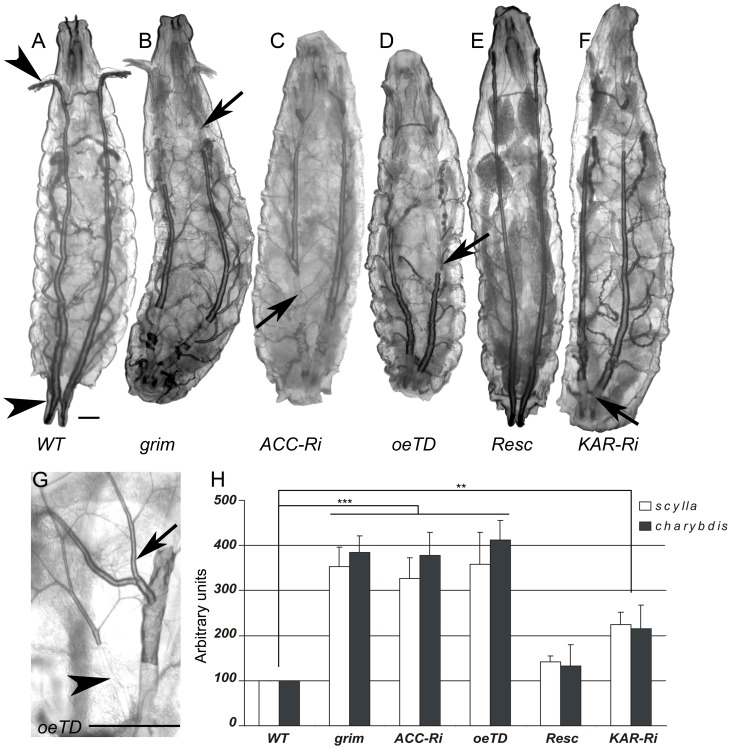
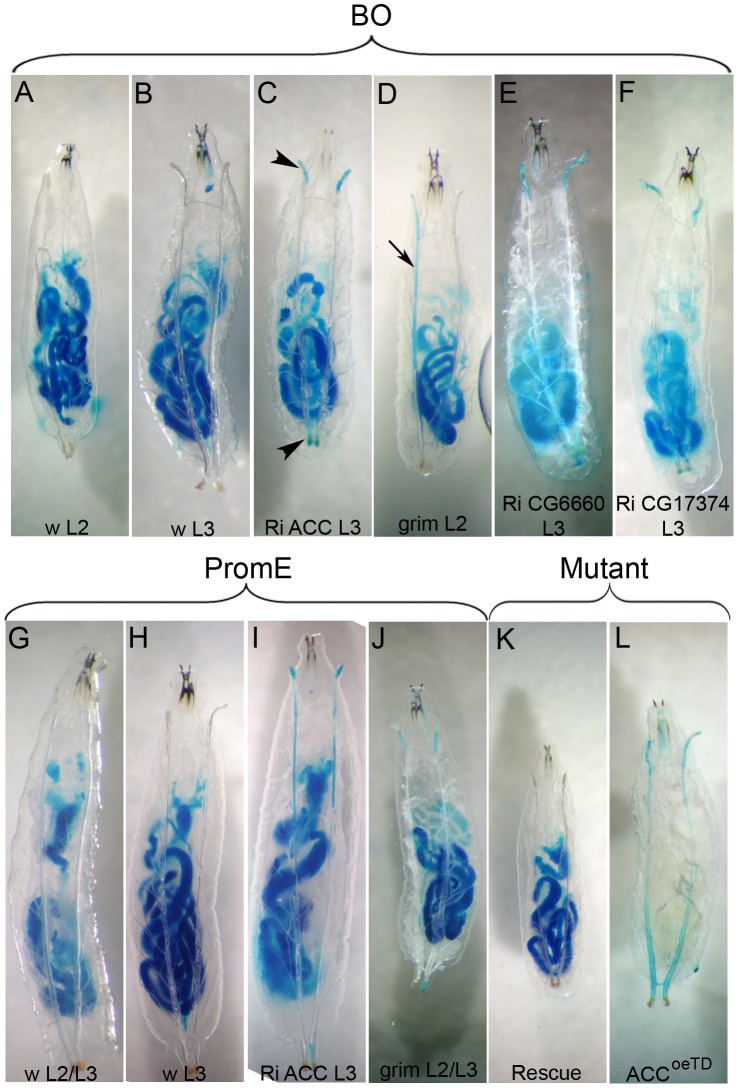
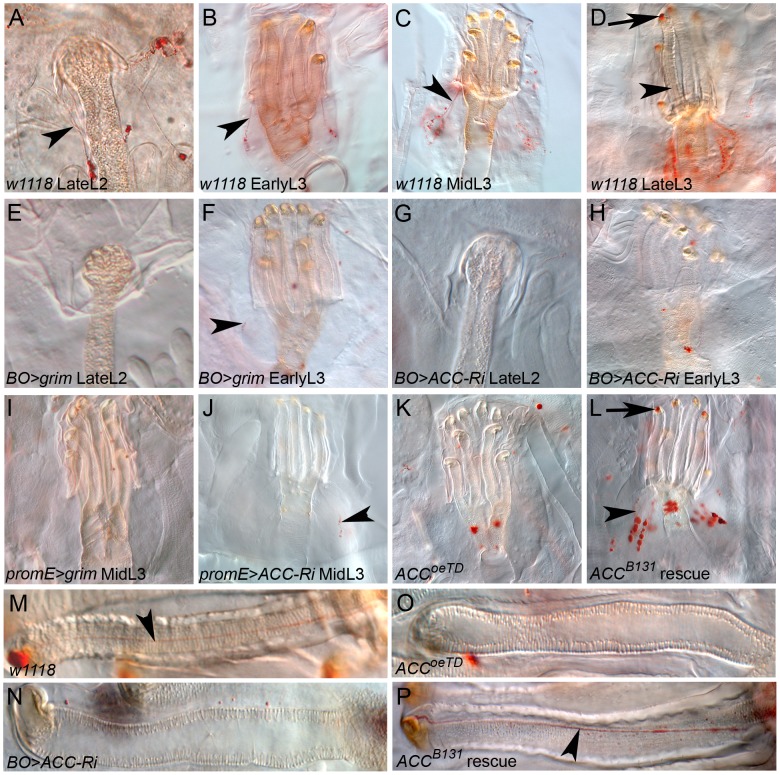
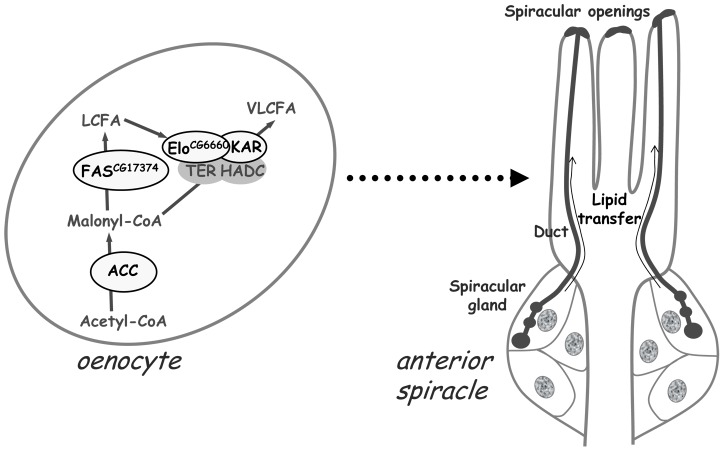
Fatty acid (FA) metabolism plays a central role in body homeostasis and related diseases. Thus, FA metabolic enzymes are attractive targets for drug therapy. Mouse studies on Acetyl-coenzymeA-carboxylase (ACC), the rate-limiting enzyme for FA synthesis, have highlighted its homeostatic role in liver and adipose tissue. We took advantage of the powerful genetics of Drosophila melanogaster to investigate the role of the unique Drosophila ACC homologue in the fat body and the oenocytes. The fat body accomplishes hepatic and storage functions, whereas the oenocytes are proposed to produce the cuticular lipids and to contribute to the hepatic function. RNA-interfering disruption of ACC in the fat body does not affect viability but does result in a dramatic reduction in triglyceride storage and a concurrent increase in glycogen accumulation. These metabolic perturbations further highlight the role of triglyceride and glycogen storage in controlling circulatory sugar levels, thereby validating Drosophila as a relevant model to explore the tissue-specific function of FA metabolic enzymes. In contrast, ACC disruption in the oenocytes through RNA-interference or tissue-targeted mutation induces lethality, as does oenocyte ablation. Surprisingly, this lethality is associated with a failure in the watertightness of the spiracles-the organs controlling the entry of air into the trachea. At the cellular level, we have observed that, in defective spiracles, lipids fail to transfer from the spiracular gland to the point of air entry. This phenotype is caused by disrupted synthesis of a putative very-long-chain-FA (VLCFA) within the oenocytes, which ultimately results in a lethal anoxic issue. Preventing liquid entry into respiratory systems is a universal issue for air-breathing animals. Here, we have shown that, in Drosophila, this process is controlled by a putative VLCFA produced within the oenocytes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The regulation of triglyceride storage by ornithine decarboxylase (Odc1) in Drosophila.Biochem Biophys Res Commun. 2020 Mar 5;523(2):429-433. doi: 10.1016/j.bbrc.2019.12.078. Epub 2019 Dec 24. Biochem Biophys Res Commun. 2020. PMID: 31870547
-
Flexible origin of hydrocarbon/pheromone precursors in Drosophila melanogaster.J Lipid Res. 2015 Nov;56(11):2094-101. doi: 10.1194/jlr.M060368. Epub 2015 Sep 9. J Lipid Res. 2015. PMID: 26353752 Free PMC article.
-
RiboTag translatomic profiling of Drosophila oenocytes under aging and induced oxidative stress.BMC Genomics. 2019 Jan 16;20(1):50. doi: 10.1186/s12864-018-5404-4. BMC Genomics. 2019. PMID: 30651069 Free PMC article.
-
The acetyl-CoA carboxylase enzyme: a target for cancer therapy?Expert Rev Anticancer Ther. 2015 Jun;15(6):667-76. doi: 10.1586/14737140.2015.1038246. Epub 2015 May 15. Expert Rev Anticancer Ther. 2015. PMID: 25979092 Review.
-
Acetyl-CoA carboxylase (ACC) as a therapeutic target for metabolic syndrome and recent developments in ACC1/2 inhibitors.Expert Opin Investig Drugs. 2019 Oct;28(10):917-930. doi: 10.1080/13543784.2019.1657825. Epub 2019 Aug 29. Expert Opin Investig Drugs. 2019. PMID: 31430206 Review.
Cited by
-
Deficiency of Acetyl-CoA Carboxylase Impairs Digestion, Lipid Synthesis, and Reproduction in the Kissing Bug Rhodnius prolixus.Front Physiol. 2022 Jul 22;13:934667. doi: 10.3389/fphys.2022.934667. eCollection 2022. Front Physiol. 2022. PMID: 35936892 Free PMC article.
-
Fat body glycolysis defects inhibit mTOR and promote distant muscle disorganization through TNF-α/egr and ImpL2 signaling in Drosophila larvae.EMBO Rep. 2024 Oct;25(10):4410-4432. doi: 10.1038/s44319-024-00241-3. Epub 2024 Sep 9. EMBO Rep. 2024. PMID: 39251827 Free PMC article.
-
Genetic Underpinnings of Cuticular Hydrocarbon Biosynthesis in the German Cockroach, Blattella germanica (L.): Progress and Perspectives.J Chem Ecol. 2024 Dec;50(12):955-968. doi: 10.1007/s10886-024-01509-7. Epub 2024 May 10. J Chem Ecol. 2024. PMID: 38727793 Review.
-
Steroid Hormone Signaling Is Essential for Pheromone Production and Oenocyte Survival.PLoS Genet. 2016 Jun 22;12(6):e1006126. doi: 10.1371/journal.pgen.1006126. eCollection 2016 Jun. PLoS Genet. 2016. PMID: 27333054 Free PMC article.
-
Fatty acid synthase cooperates with glyoxalase 1 to protect against sugar toxicity.PLoS Genet. 2015 Feb 18;11(2):e1004995. doi: 10.1371/journal.pgen.1004995. eCollection 2015 Feb. PLoS Genet. 2015. PMID: 25692475 Free PMC article.
References
-
- Iwanaga T, Tsutsumi R, Noritake J, Fukata Y, Fukata M (2009) Dynamic protein palmitoylation in cellular signaling. Prog Lipid Res 48: 117–127. - PubMed
-
- Menendez JA, Lupu R (2007) Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat Rev Cancer 7: 763–777. - PubMed
-
- Menendez JA, Vazquez-Martin A, Ortega FJ, Fernandez-Real JM (2009) Fatty acid synthase: association with insulin resistance, type 2 diabetes, and cancer. Clin Chem 55: 425–438. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
