

Deletion of the huntingtin proline-rich region does not significantly affect normal huntingtin function in mice
- PMID: 22956985
- PMCID: PMC3432989
- DOI: 10.3233/JHD-2012-120016
Deletion of the huntingtin proline-rich region does not significantly affect normal huntingtin function in mice
Abstract
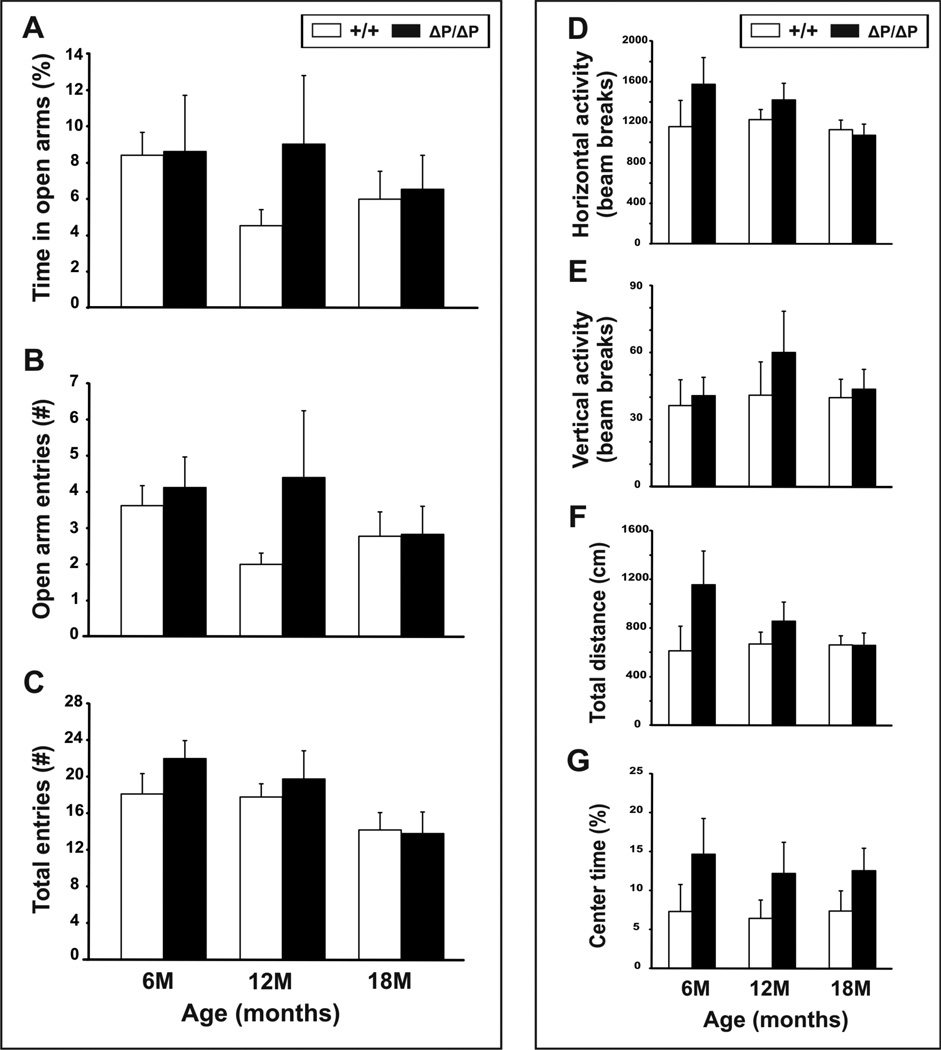
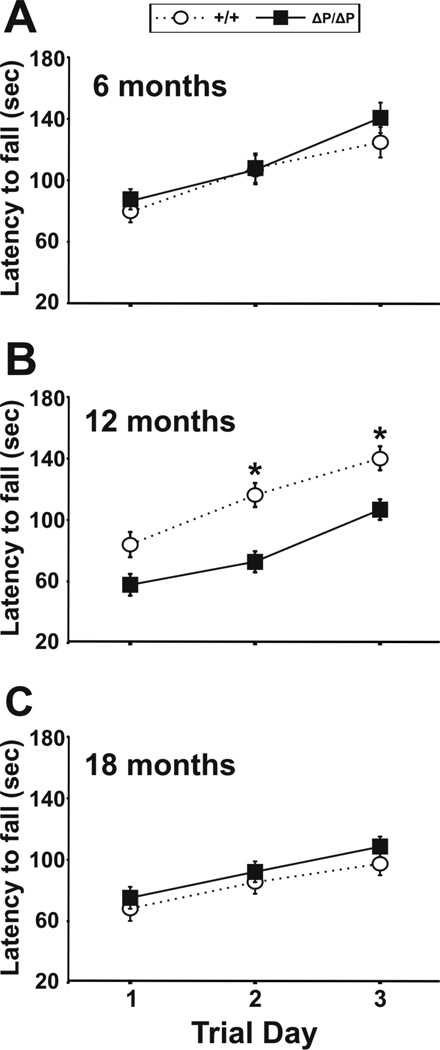
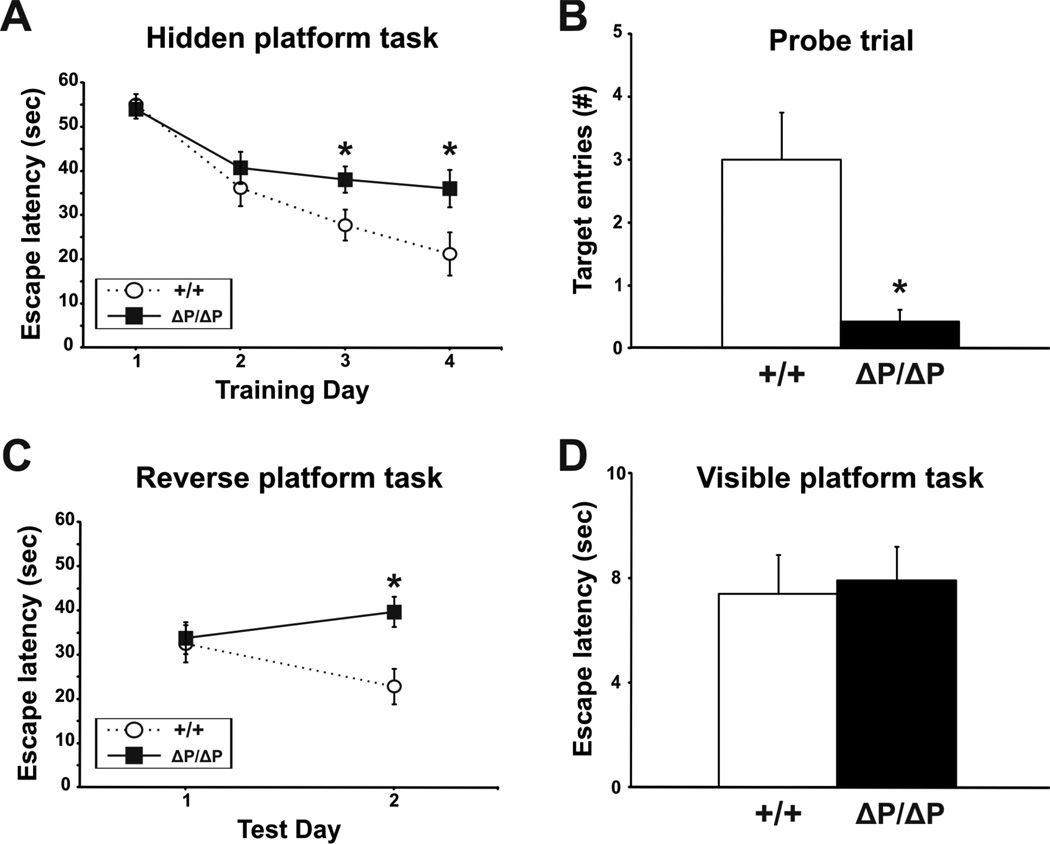
The N-terminus of Huntingtin, the protein encoded by the Huntington's disease gene, contains a stretch of polyglutamine residues that is expanded in Huntington's disease. The polyglutamine stretch is flanked by two conserved protein domains in vertebrates: an N1-17 domain, and a proline-rich region (PRR). The PRR can modulate the structure of the adjacent polyglutamine stretch, and is a binding site for several interacting proteins. To determine the role of the PRR in Huntingtin function, we have generated a knock-in allele of the mouse Huntington's disease gene homolog that expresses full-length normal huntingtin lacking the PRR. Mice that are homozygous for the huntingtin PRR deletion are born at the normal Mendelian frequency, suggesting that the PRR is not required for essential huntingtin functions during embryonic development. Moreover, adult homozygous mutants did not exhibit any significant differences from wild-type controls in general motor function and motor learning. However, 18 month-old male, but not female, homozygous PRR deletion mutants exhibited deficits in the Morris water task, suggesting that age-dependent spatial learning and memory may be affected in a sex-specific fashion by the huntingtin PRR deletion.
Conflict of interest statement
Conflict of interest statement
The authors have no conflicts of interest arising from the publication of this paper.
Figures



References
-
- Pecheux C, Gall AL, Kaplan JC, Dode C. Sequence analysis of the CAG triplet repeats region in the Huntington disease gene (IT15) in several mammalian species. Ann Genet. 1996;39(2):81–86. - PubMed
-
- Karlovich CA, John RM, Ramirez L, Stainier DY, Myers RM. Characterization of the Huntington's disease (HD) gene homologue in the zebrafish Danio rerio. Gene. 1998 Sep 14;217(1–2):117–125. - PubMed
-
- Baxendale S, Abdulla S, Elgar G, Buck D, Berks M, Micklem G, Durbin R, Bates G, Brenner S, Beck S. Comparative sequence analysis of the human and pufferfish Huntington's disease genes. Nat Genet. 1995 May;10(1):67–76. - PubMed
-
- Li Z, Karlovich CA, Fish MP, Scott MP, Myers RM. A putative Drosophila homolog of the Huntington's disease gene. Hum Mol Genet. 1999 Sep;8(9):1807–1815. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
