

mTOR complex 1 plays critical roles in hematopoiesis and Pten-loss-evoked leukemogenesis
- PMID: 22958934
- PMCID: PMC3743253
- DOI: 10.1016/j.stem.2012.06.009
mTOR complex 1 plays critical roles in hematopoiesis and Pten-loss-evoked leukemogenesis
Abstract
The mechanistic target of rapamycin (mTOR) pathway serves as a key sensor of cellular-energetic state and functions to maintain tissue homeostasis. Hyperactivation of the mTOR pathway impairs hematopoietic stem cell (HSC) function and is associated with leukemogenesis. However, the roles of the unique mTOR complexes (mTORCs) in hematopoiesis and leukemogenesis have not been adequately elucidated. We deleted the mTORC1 component, regulatory-associated protein of mTOR (Raptor), in mouse HSCs and its loss causes a nonlethal phenotype characterized by pancytopenia, splenomegaly, and the accumulation of monocytoid cells. Furthermore, Raptor is required for HSC regeneration, and plays largely nonredundant roles with rapamycin-insensitive companion of mTOR (Rictor) in these processes. Ablation of Raptor also significantly extends survival of mice in models of leukemogenesis evoked by Pten deficiency. These data delineate critical roles for mTORC1 in hematopoietic function and leukemogenesis and inform clinical strategies based on chronic mTORC1 inhibition.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
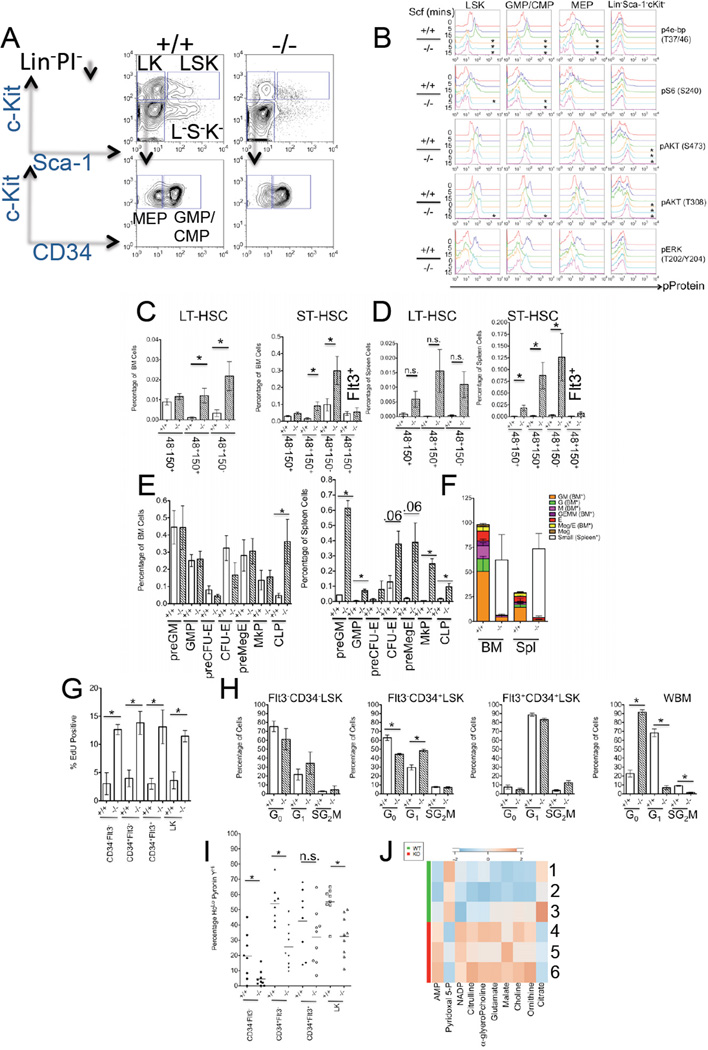
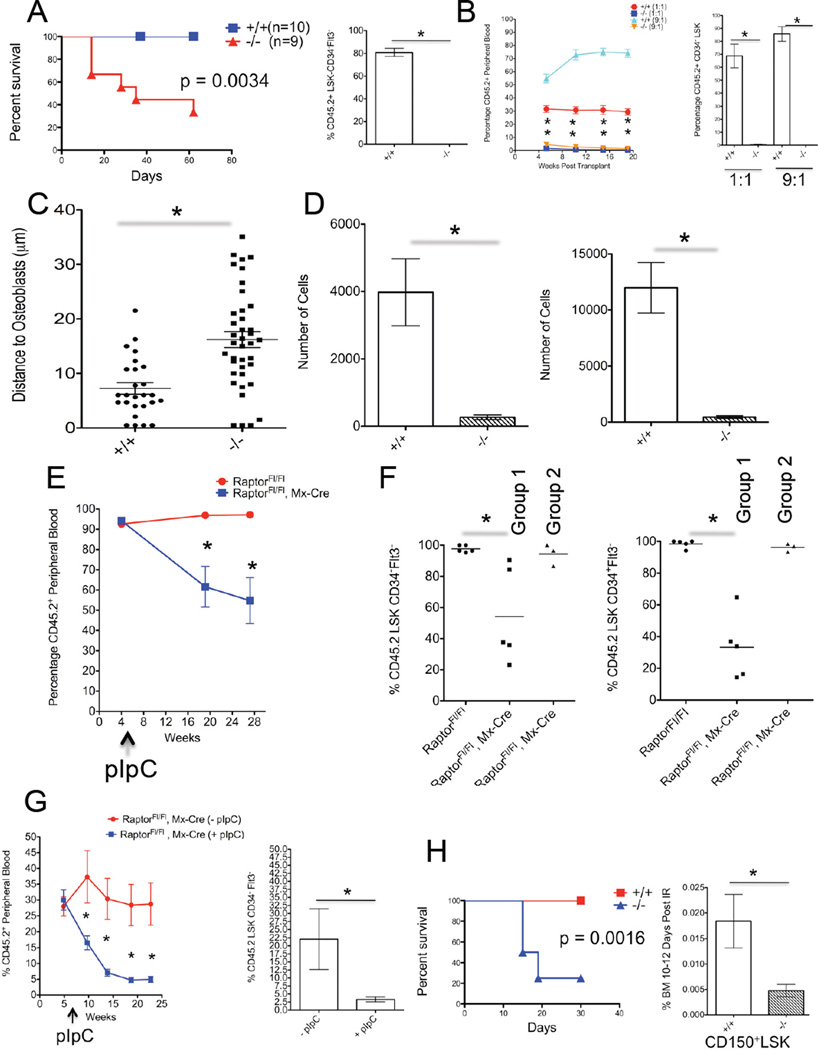
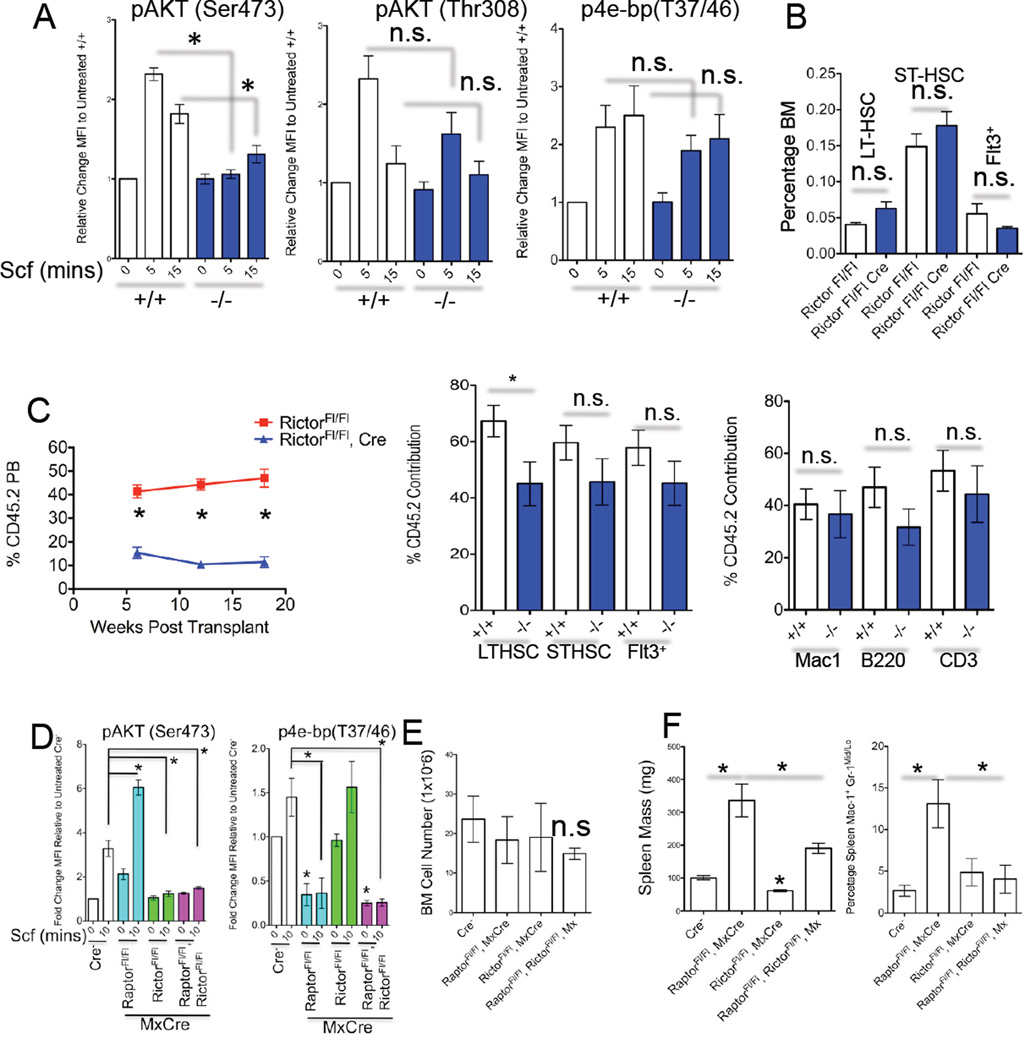
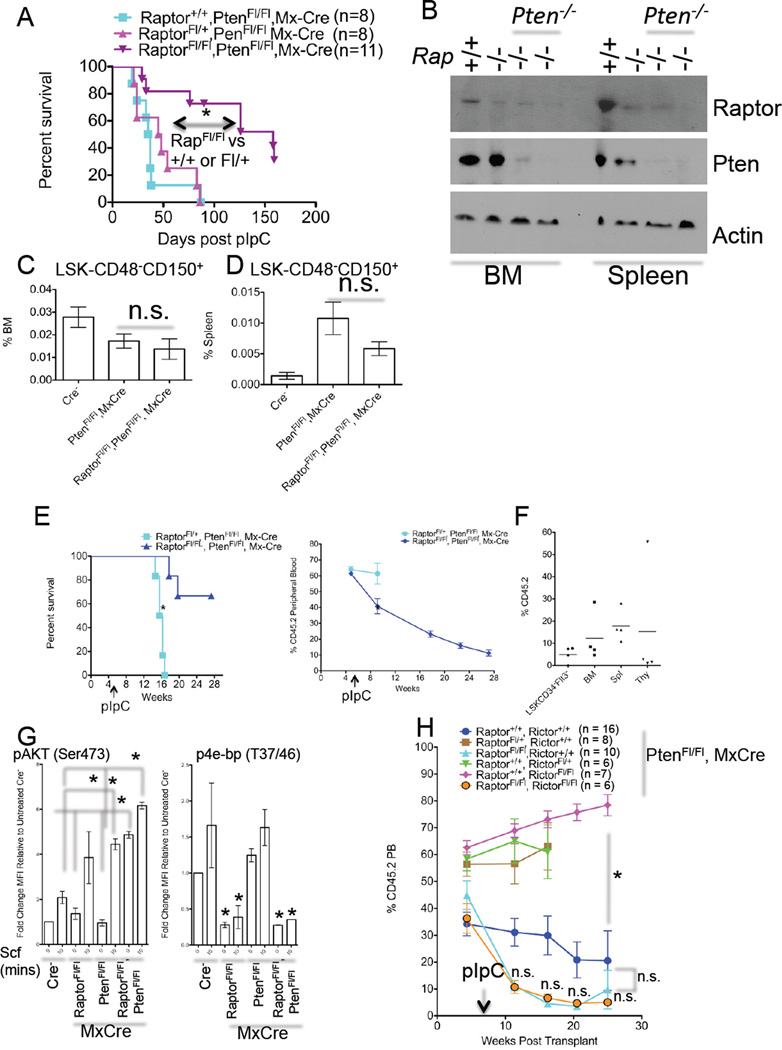
Comment in
-
Distinct and dynamic requirements for mTOR signaling in hematopoiesis and leukemogenesis.Cell Stem Cell. 2012 Sep 7;11(3):281-2. doi: 10.1016/j.stem.2012.08.007. Cell Stem Cell. 2012. PMID: 22958924
References
-
- Carracedo A, Pandolfi PP. The PTEN-PI3K pathway: of feedbacks and cross-talks. Oncogene. 2008;27:5527–5541. - PubMed
-
- Chapuis N, Tamburini J, Green AS, Willems L, Bardet V, Park S, Lacombe C, Mayeux P, Bouscary D. Perspectives on inhibiting mTOR as a future treatment strategy for hematological malignancies. Leukemia. 2010;23:1686–1699. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- U01 CA105423/CA/NCI NIH HHS/United States
- R01 HL044851/HL/NHLBI NIH HHS/United States
- K01DK092300/DK/NIDDK NIH HHS/United States
- HL097748/HL/NHLBI NIH HHS/United States
- K99 CA158461/CA/NCI NIH HHS/United States
- R00 CA129613/CA/NCI NIH HHS/United States
- P01 CA066996/CA/NCI NIH HHS/United States
- U01 HL100402/HL/NHLBI NIH HHS/United States
- HL097794/HL/NHLBI NIH HHS/United States
- K01 DK092300/DK/NIDDK NIH HHS/United States
- R01 HL097794/HL/NHLBI NIH HHS/United States
- R01 HL097748/HL/NHLBI NIH HHS/United States
- R01 DK050234/DK/NIDDK NIH HHS/United States
- DK050234/DK/NIDDK NIH HHS/United States
- CA66996/CA/NCI NIH HHS/United States
- R00 CA158461/CA/NCI NIH HHS/United States
- DK049216/DK/NIDDK NIH HHS/United States
- HL100402/HL/NHLBI NIH HHS/United States
- CA105423/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
