

How are inner hair cells stimulated? Evidence for multiple mechanical drives
- PMID: 22959529
- PMCID: PMC3549570
- DOI: 10.1016/j.heares.2012.08.005
How are inner hair cells stimulated? Evidence for multiple mechanical drives
Abstract
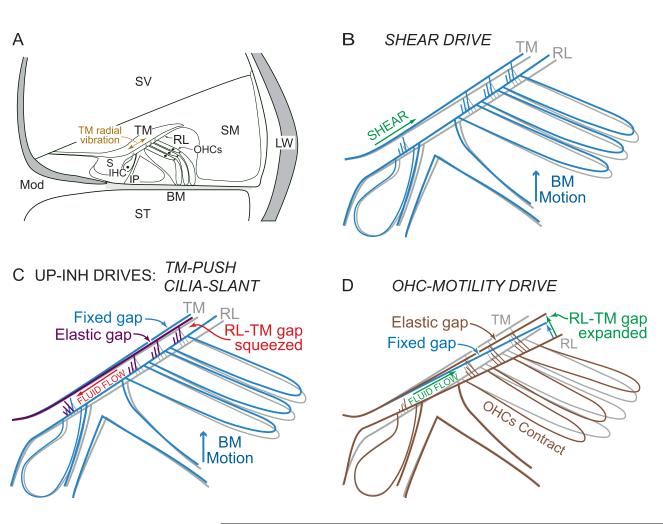
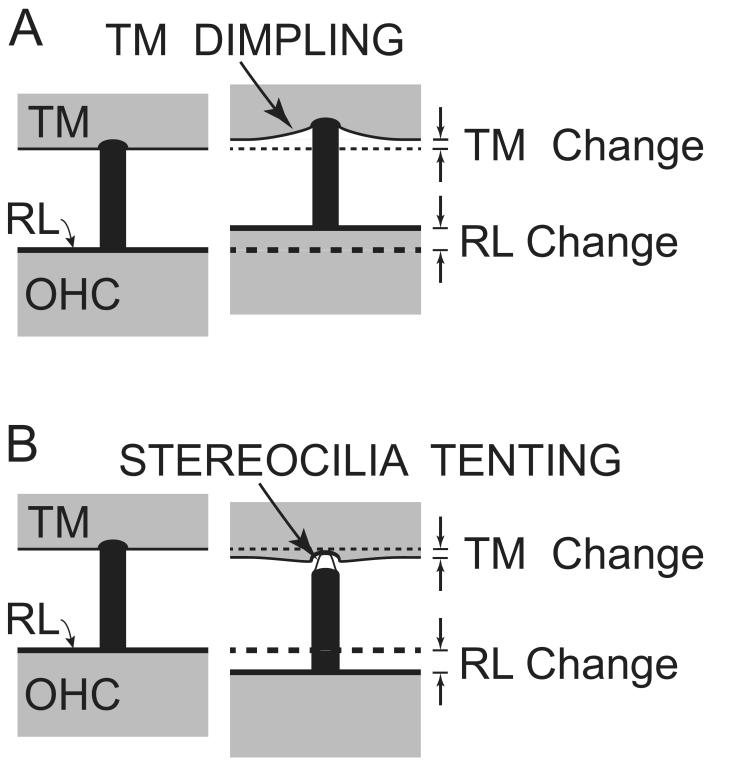
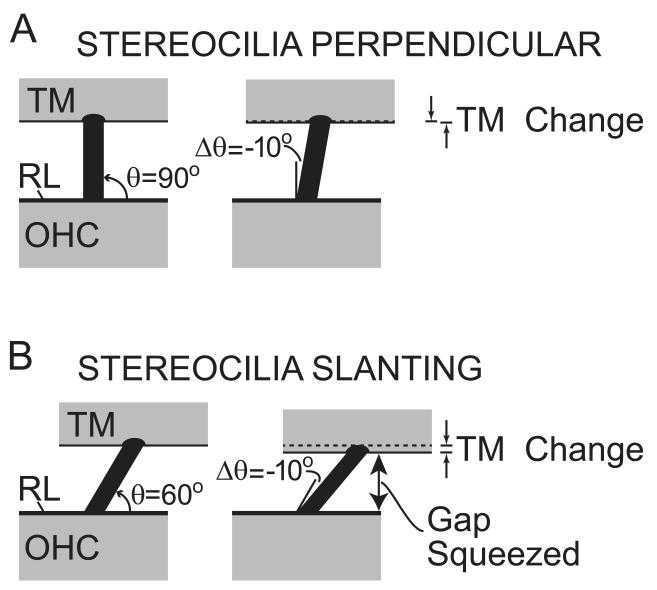
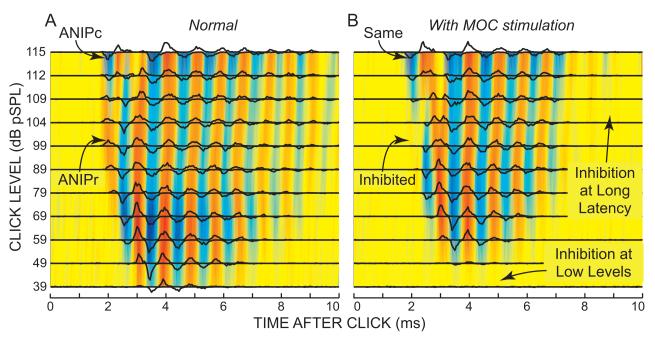
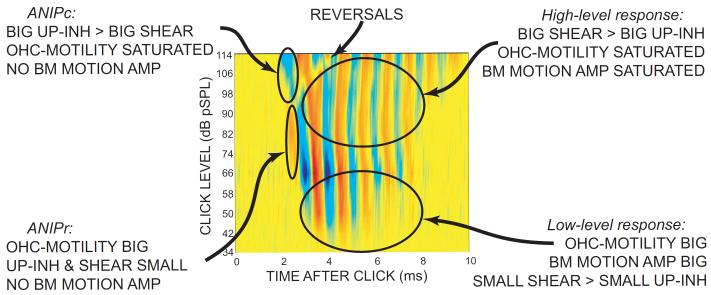
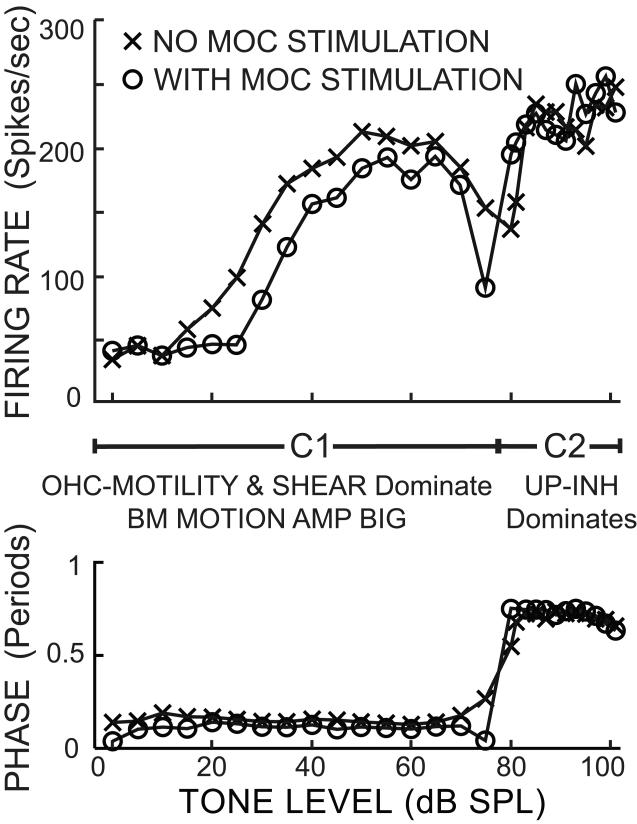
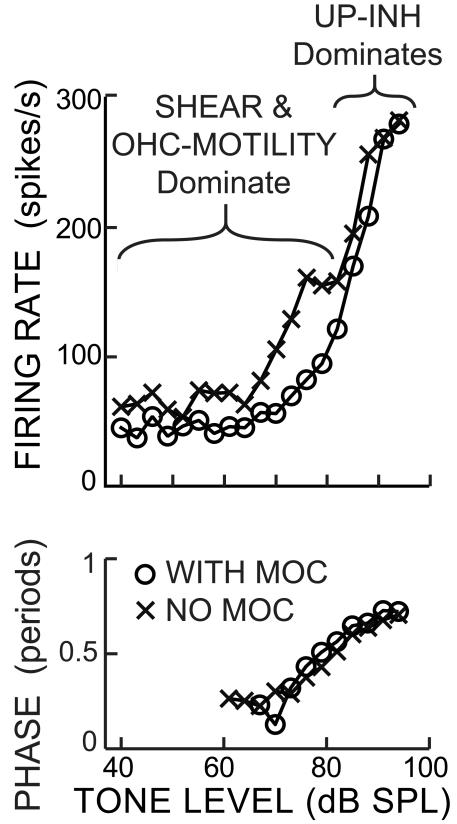
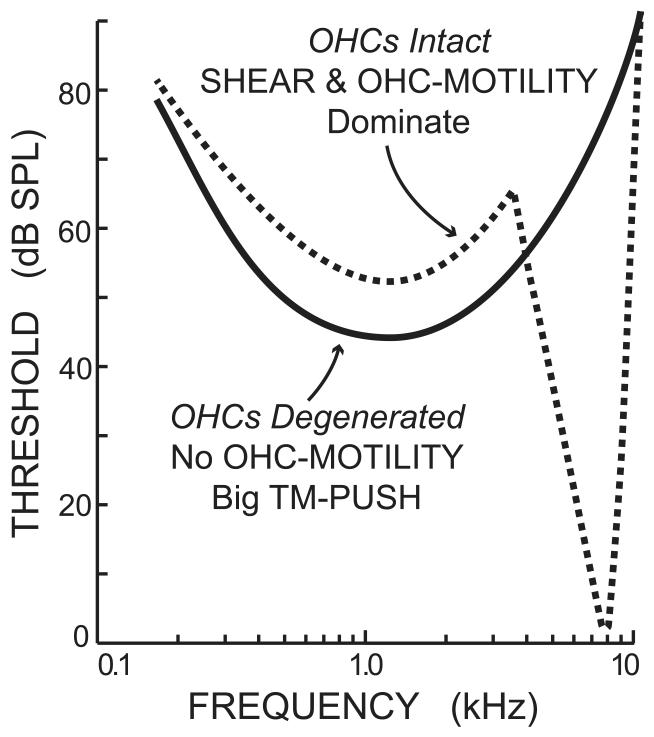
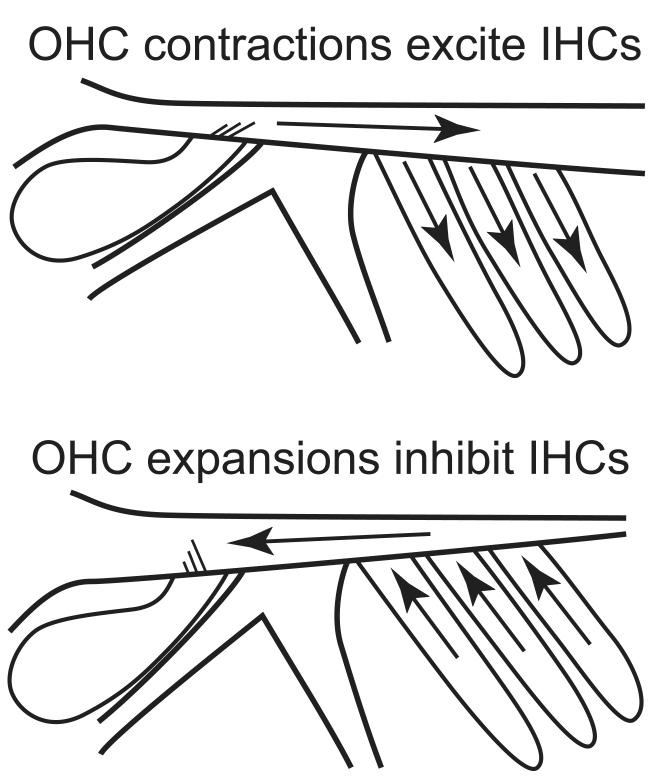
Recent studies indicate that the gap over outer hair cells (OHCs) between the reticular lamina (RL) and the tectorial membrane (TM) varies cyclically during low-frequency sounds. Variation in the RL-TM gap produces radial fluid flow in the gap that can drive inner hair cell (IHC) stereocilia. Analysis of RL-TM gap changes reveals three IHC drives in addition to classic SHEAR. For upward basilar-membrane (BM) motion, IHC stereocilia are deflected in the excitatory direction by SHEAR and OHC-MOTILITY, but in the inhibitory direction by TM-PUSH and CILIA-SLANT. Upward BM motion causes OHC somatic contraction which tilts the RL, compresses the RL-TM gap over IHCs and expands the RL-TM gap over OHCs, thereby producing an outward (away from the IHCs) radial fluid flow which is the OHC-MOTILITY drive. For upward BM motion, the force that moves the TM upward also compresses the RL-TM gap over OHCs causing inward radial flow past IHCs which is the TM-PUSH drive. Motions that produce large tilting of OHC stereocilia squeeze the supra-OHC RL-TM gap and caused inward radial flow past IHCs which is the CILIA-SLANT drive. Combinations of these drives explain: (1) the reversal at high sound levels of auditory nerve (AN) initial peak (ANIP) responses to clicks, and medial olivocochlear (MOC) inhibition of ANIP responses below, but not above, the ANIP reversal, (2) dips and phase reversals in AN responses to tones in cats and chinchillas, (3) hypersensitivity and phase reversals in tuning-curve tails after OHC ablation, and (4) MOC inhibition of tail-frequency AN responses. The OHC-MOTILITY drive provides another mechanism, in addition to BM motion amplification, that uses active processes to enhance the output of the cochlea. The ability of these IHC drives to explain previously anomalous data provides strong, although indirect, evidence that these drives are significant and presents a new view of how the cochlea works at frequencies below 3 kHz.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Allen JB. Cochlear micromechanics--a physical model of transduction. J Acoust Soc Am. 1980;68:1660–70. - PubMed
-
- Baumgart J, Chiaradia C, Fleischer M, Yarin Y, Grundmann R, Gummer AW. Fluid mechanics in the subtectorial space. In: Cooper NP, Kemp DT, editors. Concepts and Challenges in the Biophysics of Hearing. World Scientific; Singapore: 2009. pp. 288–293.
-
- Cheatham MA, Dallos P. The level dependence of response phase: observations from cochlear hair cells. J Acoust Soc Am. 1998;104:356–69. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
