

Activity-dependent human brain coding/noncoding gene regulatory networks
- PMID: 22960213
- PMCID: PMC3522156
- DOI: 10.1534/genetics.112.145128
Activity-dependent human brain coding/noncoding gene regulatory networks
Abstract
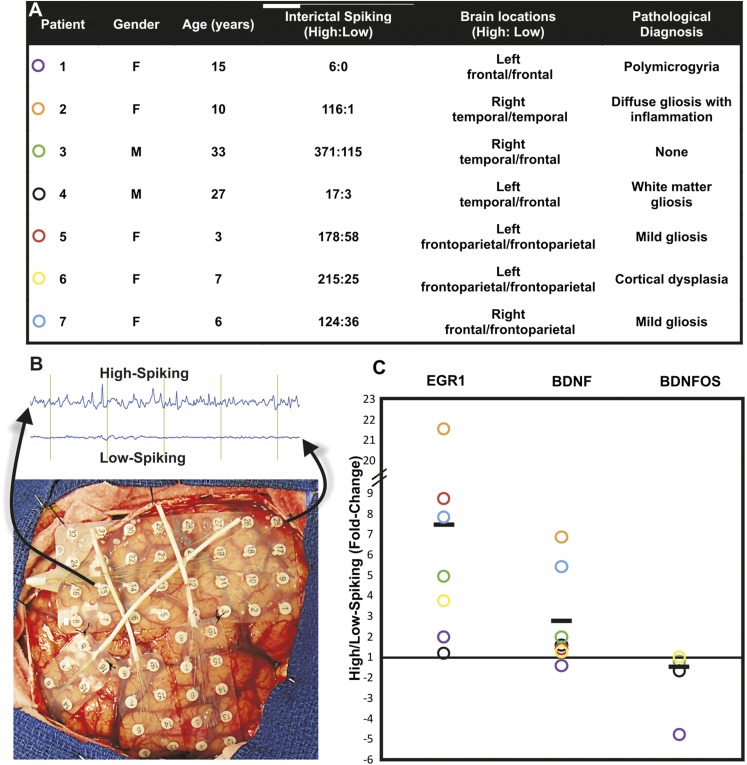
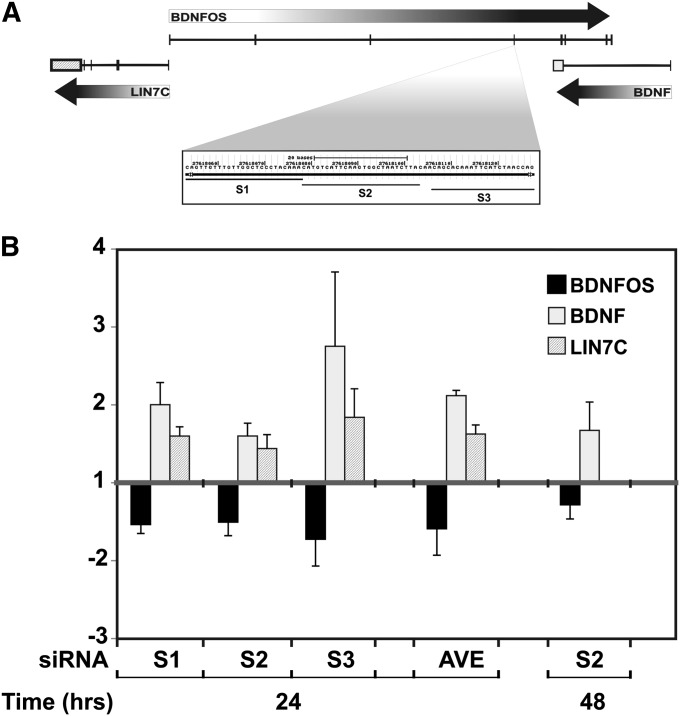
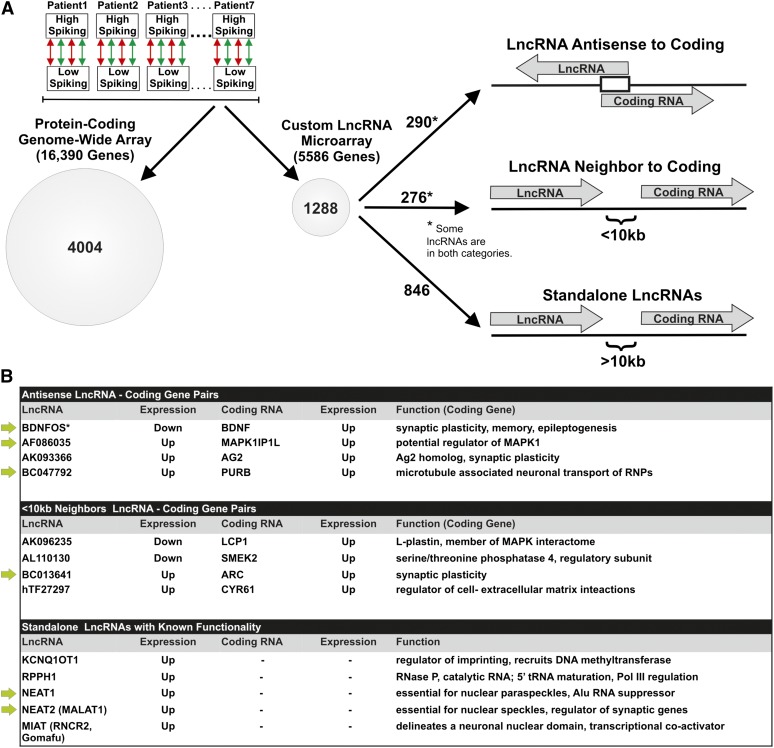
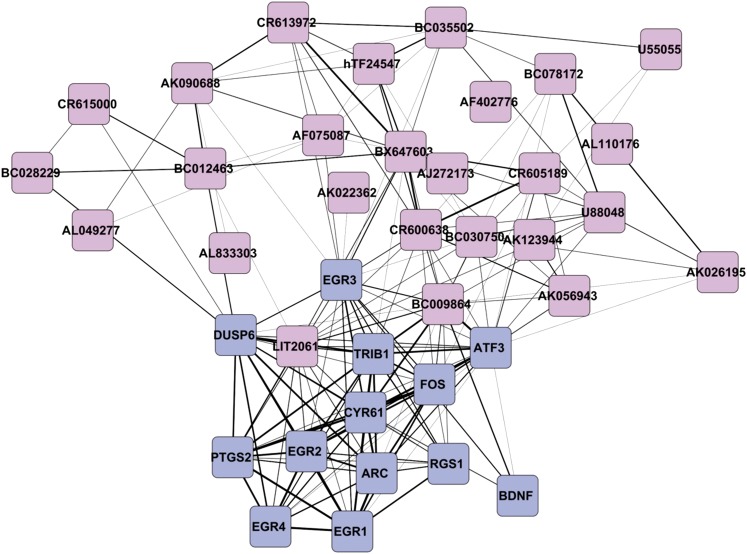
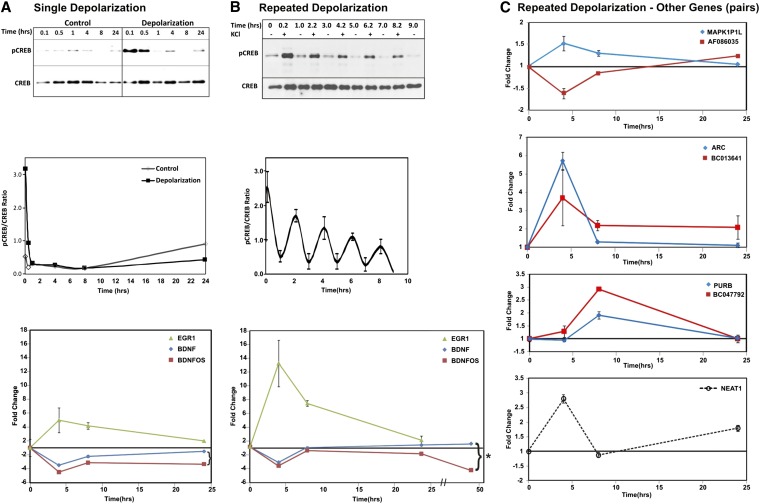
While most gene transcription yields RNA transcripts that code for proteins, a sizable proportion of the genome generates RNA transcripts that do not code for proteins, but may have important regulatory functions. The brain-derived neurotrophic factor (BDNF) gene, a key regulator of neuronal activity, is overlapped by a primate-specific, antisense long noncoding RNA (lncRNA) called BDNFOS. We demonstrate reciprocal patterns of BDNF and BDNFOS transcription in highly active regions of human neocortex removed as a treatment for intractable seizures. A genome-wide analysis of activity-dependent coding and noncoding human transcription using a custom lncRNA microarray identified 1288 differentially expressed lncRNAs, of which 26 had expression profiles that matched activity-dependent coding genes and an additional 8 were adjacent to or overlapping with differentially expressed protein-coding genes. The functions of most of these protein-coding partner genes, such as ARC, include long-term potentiation, synaptic activity, and memory. The nuclear lncRNAs NEAT1, MALAT1, and RPPH1, composing an RNAse P-dependent lncRNA-maturation pathway, were also upregulated. As a means to replicate human neuronal activity, repeated depolarization of SY5Y cells resulted in sustained CREB activation and produced an inverse pattern of BDNF-BDNFOS co-expression that was not achieved with a single depolarization. RNAi-mediated knockdown of BDNFOS in human SY5Y cells increased BDNF expression, suggesting that BDNFOS directly downregulates BDNF. Temporal expression patterns of other lncRNA-messenger RNA pairs validated the effect of chronic neuronal activity on the transcriptome and implied various lncRNA regulatory mechanisms. lncRNAs, some of which are unique to primates, thus appear to have potentially important regulatory roles in activity-dependent human brain plasticity.
Figures
Similar articles
-
Differential long noncoding RNA/mRNA expression profiling and functional network analysis during osteogenic differentiation of human bone marrow mesenchymal stem cells.Stem Cell Res Ther. 2017 Feb 7;8(1):30. doi: 10.1186/s13287-017-0485-6. Stem Cell Res Ther. 2017. PMID: 28173844 Free PMC article.
-
Decoding the Long Noncoding RNA During Cardiac Maturation: A Roadmap for Functional Discovery.Circ Cardiovasc Genet. 2016 Oct;9(5):395-407. doi: 10.1161/CIRCGENETICS.115.001363. Epub 2016 Sep 2. Circ Cardiovasc Genet. 2016. PMID: 27591185 Free PMC article.
-
Microarray Profiling and Co-Expression Network Analysis of LncRNAs and mRNAs in Neonatal Rats Following Hypoxic-ischemic Brain Damage.Sci Rep. 2015 Sep 9;5:13850. doi: 10.1038/srep13850. Sci Rep. 2015. PMID: 26349411 Free PMC article.
-
Long non-coding RNAs: From disease code to drug role.Acta Pharm Sin B. 2021 Feb;11(2):340-354. doi: 10.1016/j.apsb.2020.10.001. Epub 2020 Oct 10. Acta Pharm Sin B. 2021. PMID: 33643816 Free PMC article. Review.
-
Roles of long noncoding RNAs in brain development, functional diversification and neurodegenerative diseases.Brain Res Bull. 2013 Aug;97:69-80. doi: 10.1016/j.brainresbull.2013.06.001. Epub 2013 Jun 10. Brain Res Bull. 2013. PMID: 23756188 Review.
Cited by
-
Role of Long Noncoding RNAs in Parkinson's Disease: Putative Biomarkers and Therapeutic Targets.Parkinsons Dis. 2020 Jun 12;2020:5374307. doi: 10.1155/2020/5374307. eCollection 2020. Parkinsons Dis. 2020. PMID: 32617144 Free PMC article. Review.
-
Challenges in LncRNA Biology: Views and Opinions.Noncoding RNA. 2024 Aug 1;10(4):43. doi: 10.3390/ncrna10040043. Noncoding RNA. 2024. PMID: 39195572 Free PMC article.
-
Long non-coding RNAs: novel targets for nervous system disease diagnosis and therapy.Neurotherapeutics. 2013 Oct;10(4):632-46. doi: 10.1007/s13311-013-0199-0. Neurotherapeutics. 2013. PMID: 23817781 Free PMC article. Review.
-
Sense-antisense gene pairs: sequence, transcription, and structure are not conserved between human and mouse.Front Genet. 2013 Sep 26;4:183. doi: 10.3389/fgene.2013.00183. eCollection 2013. Front Genet. 2013. PMID: 24133500 Free PMC article.
-
Value of long non-coding RNA Rpph1 in esophageal cancer and its effect on cancer cell sensitivity to radiotherapy.World J Gastroenterol. 2020 Apr 21;26(15):1775-1791. doi: 10.3748/wjg.v26.i15.1775. World J Gastroenterol. 2020. PMID: 32351293 Free PMC article.
References
-
- Bates D., Maechler M., Bolker B., 2009. lme4: Linear mexed-effects models using S4 classes. Available at: https://r-forge.r-project.org/R/?group_id=60. Accessed: September, 2012.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
