

Capping protein modulates the dynamic behavior of actin filaments in response to phosphatidic acid in Arabidopsis
- PMID: 22960908
- PMCID: PMC3480299
- DOI: 10.1105/tpc.112.103945
Capping protein modulates the dynamic behavior of actin filaments in response to phosphatidic acid in Arabidopsis
Abstract
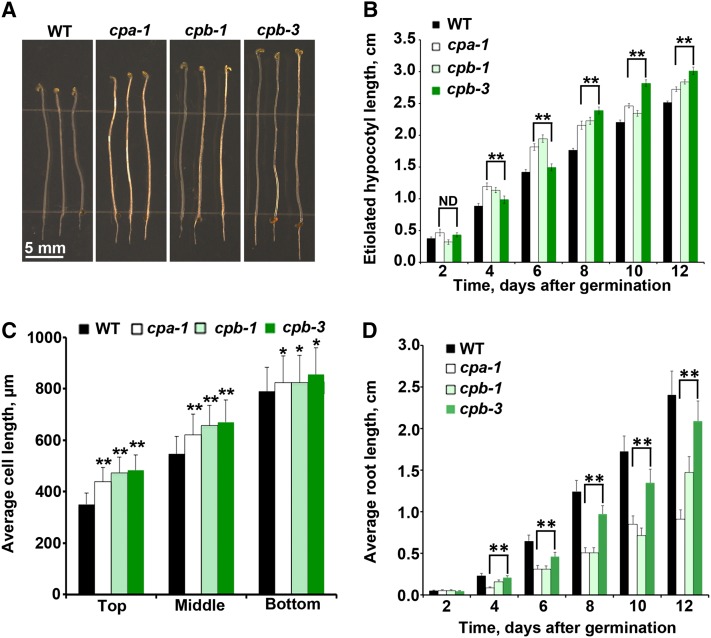
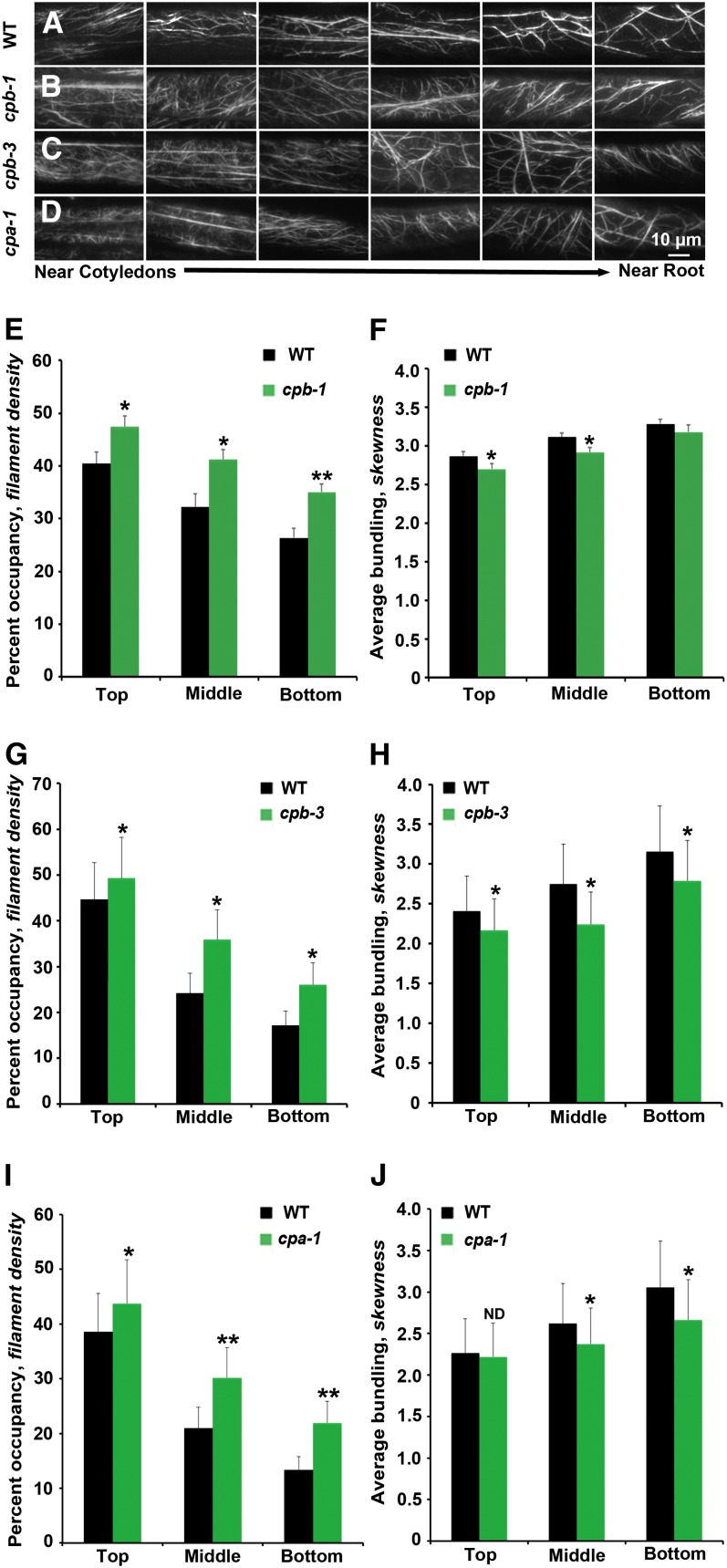
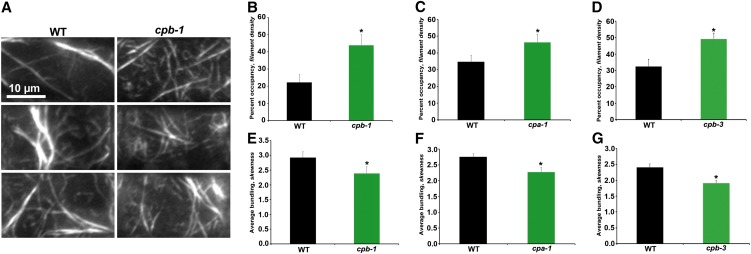
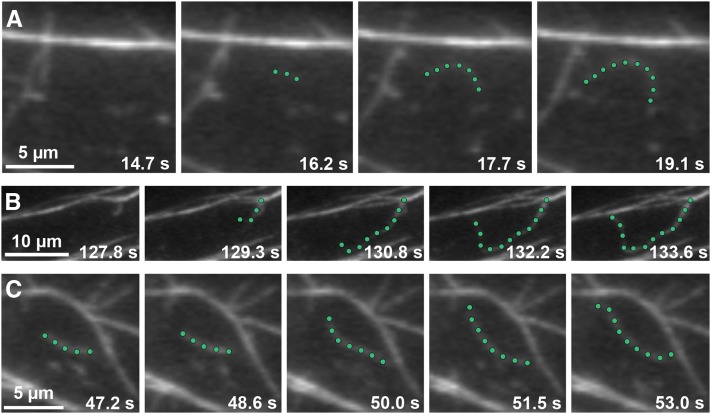
Remodeling of actin filament arrays in response to biotic and abiotic stimuli is thought to require precise control over the generation and availability of filament ends. Heterodimeric capping protein (CP) is an abundant filament capper, and its activity is inhibited by membrane signaling phospholipids in vitro. How exactly CP modulates the properties of filament ends in cells and whether its activity is coordinated by phospholipids in vivo is not well understood. By observing directly the dynamic behavior of individual filament ends in the cortical array of living Arabidopsis thaliana epidermal cells, we dissected the contribution of CP to actin organization and dynamics in response to the signaling phospholipid, phosphatidic acid (PA). Here, we examined three cp knockdown mutants and found that reduced CP levels resulted in more dynamic activity at filament ends, and this significantly enhanced filament-filament annealing and filament elongation from free ends. The cp mutants also exhibited more dense actin filament arrays. Treatment of wild-type cells with exogenous PA phenocopied the actin-based defects in cp mutants, with an increase in the density of filament arrays and enhanced annealing frequency. These cytoskeletal responses to exogenous PA were completely abrogated in cp mutants. Our data provide compelling genetic evidence that the end-capping activity of CP is inhibited by membrane signaling lipids in eukaryotic cells. Specifically, CP acts as a PA biosensor and key transducer of fluxes in membrane signaling phospholipids into changes in actin cytoskeleton dynamics.
Figures
Similar articles
-
Heterodimeric capping protein from Arabidopsis is regulated by phosphatidic acid.Mol Biol Cell. 2006 Apr;17(4):1946-58. doi: 10.1091/mbc.e05-09-0840. Epub 2006 Jan 25. Mol Biol Cell. 2006. PMID: 16436516 Free PMC article.
-
Heterodimeric capping protein from Arabidopsis is a membrane-associated, actin-binding protein.Plant Physiol. 2014 Nov;166(3):1312-28. doi: 10.1104/pp.114.242487. Epub 2014 Sep 8. Plant Physiol. 2014. PMID: 25201878 Free PMC article.
-
Arabidopsis capping protein senses cellular phosphatidic acid levels and transduces these into changes in actin cytoskeleton dynamics.Plant Signal Behav. 2012 Dec;7(12):1727-30. doi: 10.4161/psb.22472. Epub 2012 Oct 16. Plant Signal Behav. 2012. PMID: 23072985 Free PMC article.
-
Capping protein regulators fine-tune actin assembly dynamics.Nat Rev Mol Cell Biol. 2014 Oct;15(10):677-89. doi: 10.1038/nrm3869. Epub 2014 Sep 10. Nat Rev Mol Cell Biol. 2014. PMID: 25207437 Free PMC article. Review.
-
Actin dynamics in the cortical array of plant cells.Curr Opin Plant Biol. 2013 Dec;16(6):678-87. doi: 10.1016/j.pbi.2013.10.012. Epub 2013 Nov 15. Curr Opin Plant Biol. 2013. PMID: 24246228 Review.
Cited by
-
Capping proteins regulate fungal development, DON-toxisome formation and virulence in Fusarium graminearum.Mol Plant Pathol. 2020 Feb;21(2):173-187. doi: 10.1111/mpp.12887. Epub 2019 Nov 6. Mol Plant Pathol. 2020. PMID: 31693278 Free PMC article.
-
When fat is not bad: the regulation of actin dynamics by phospholipid signaling molecules.Front Plant Sci. 2014 Jan 23;5:5. doi: 10.3389/fpls.2014.00005. eCollection 2014. Front Plant Sci. 2014. PMID: 24478785 Free PMC article. Review.
-
Phospholipids: molecules regulating cytoskeletal organization in plant abiotic stress tolerance.Plant Signal Behav. 2014;9(3):e28337. doi: 10.4161/psb.28337. Epub 2014 Jan 1. Plant Signal Behav. 2014. PMID: 24589893 Free PMC article. Review.
-
Spaceflight induces novel regulatory responses in Arabidopsis seedling as revealed by combined proteomic and transcriptomic analyses.BMC Plant Biol. 2020 May 27;20(1):237. doi: 10.1186/s12870-020-02392-6. BMC Plant Biol. 2020. PMID: 32460700 Free PMC article.
-
Regulation of developmental and environmental signaling by interaction between microtubules and membranes in plant cells.Protein Cell. 2016 Feb;7(2):81-8. doi: 10.1007/s13238-015-0233-6. Epub 2015 Dec 19. Protein Cell. 2016. PMID: 26687389 Free PMC article. Review.
References
-
- Andrianantoandro E., Blanchoin L., Sept D., McCammon J.A., Pollard T.D. (2001). Kinetic mechanism of end-to-end annealing of actin filaments. J. Mol. Biol. 312: 721–730 - PubMed
-
- Bao C., Wang J., Zhang R., Zhang B., Zhang H., Zhou Y., Huang S. (May 5, 2012). Arabidopsis VILLIN2 and VILLIN3 act redundantly in sclerenchyma development via bundling of actin filaments. Plant J. http://dx.doi.org/.10.1111/j.1365-1313X.2012.05044.x - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
