

Phrenic long-term facilitation after acute intermittent hypoxia requires spinal ERK activation but not TrkB synthesis
- PMID: 22961271
- PMCID: PMC3472488
- DOI: 10.1152/japplphysiol.00098.2012
Phrenic long-term facilitation after acute intermittent hypoxia requires spinal ERK activation but not TrkB synthesis
Abstract
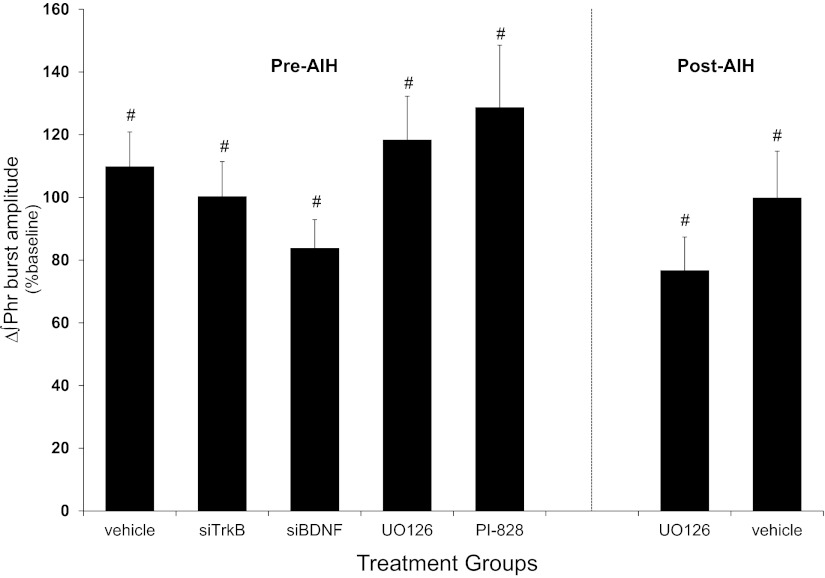
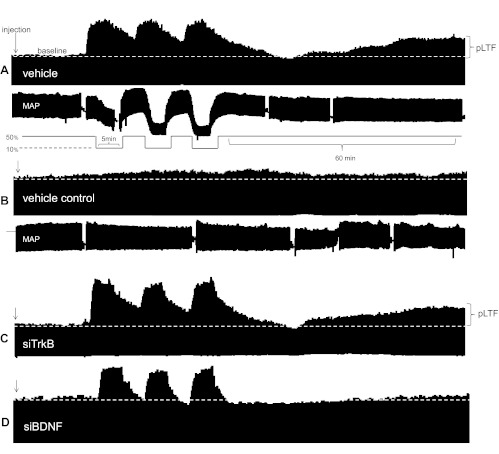
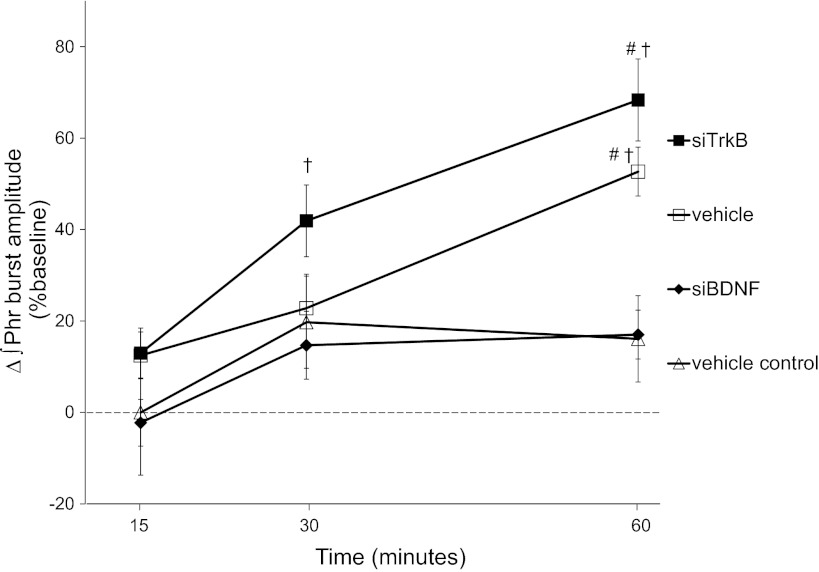
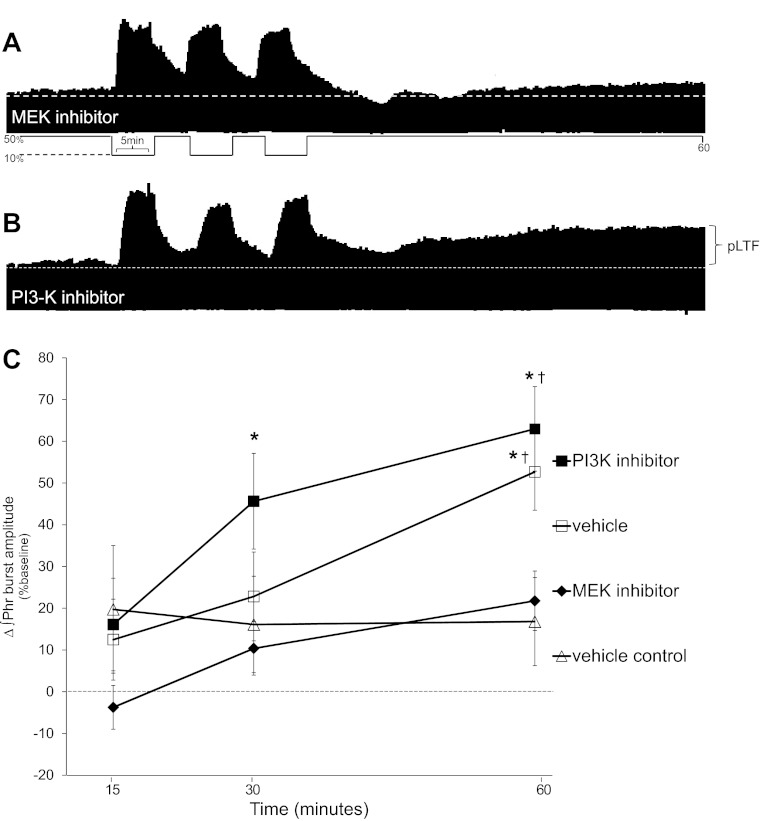
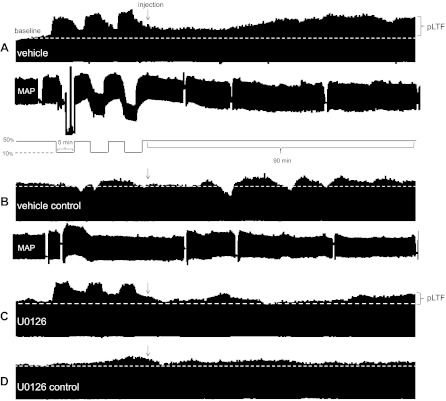
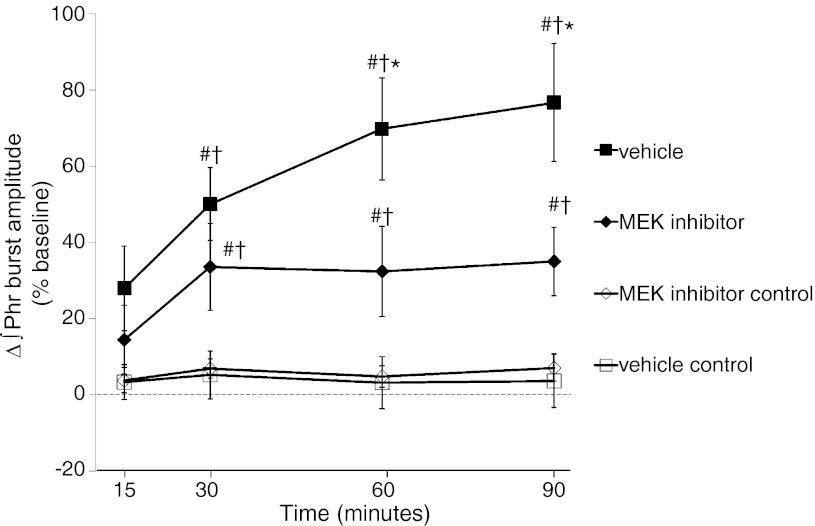
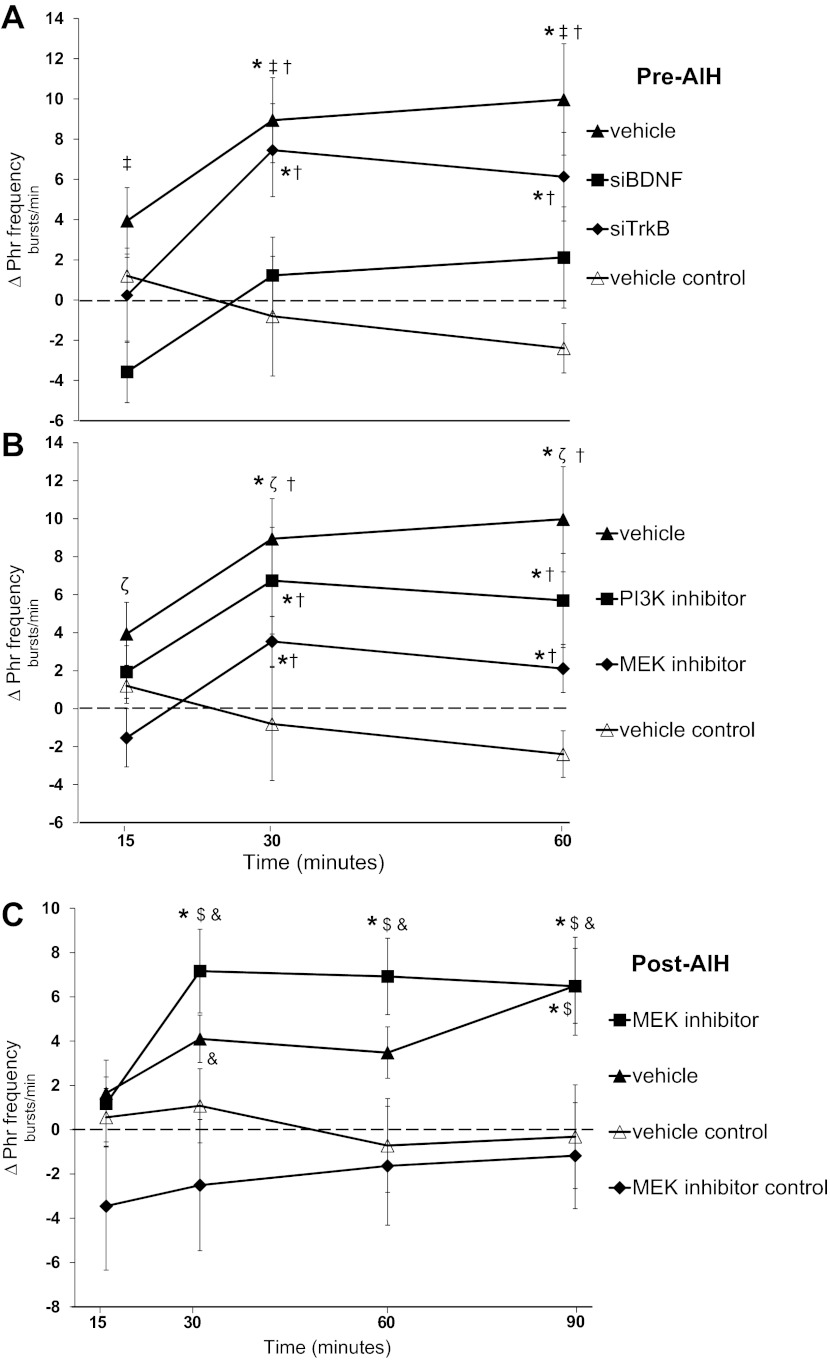
Acute intermittent hypoxia (AIH) elicits a form of spinal respiratory plasticity known as phrenic long-term facilitation (pLTF). pLTF requires spinal serotonin receptor-2 activation, the synthesis of new brain-derived neurotrophic factor (BDNF), and the activation of its high-affinity receptor tyrosine kinase, TrkB. Spinal adenosine 2A receptor activation elicits a distinct pathway to phrenic motor facilitation (pMF); this BDNF synthesis-independent pathway instead requires new synthesis of an immature TrkB isoform. Since hypoxia increases extracellular adenosine levels, we tested the hypothesis that new synthesis of TrkB and BDNF contribute to AIH-induced pLTF. Furthermore, given that signaling mechanisms "downstream" from TrkB are unknown in either mechanism, we tested the hypothesis that pLTF requires MEK/ERK and/or phosphatidylinositol 3-kinase (PI3K)/Akt activation. In anesthetized Sprague-Dawley rats, an intrathecal catheter at cervical level 4 was used to deliver drugs near the phrenic motor nucleus. Since pLTF was blocked by spinal injections of small interfering RNAs targeting BDNF mRNA but not TrkB mRNA, only new BDNF synthesis is required for AIH-induced pLTF. Pretreatment with a MEK inhibitor (U0126) blocked pLTF, whereas a PI3K inhibitor (PI-828) had no effect. Thus, AIH-induced pLTF requires MEK/ERK (not PI3K/AKT) signaling pathways. When U0126 was injected post-AIH, pLTF development was halted but not reversed, suggesting that ERK is critical for the development but not maintenance of pLTF. Thus, there are clear mechanistic distinctions between AIH-induced pLTF (i.e., BDNF synthesis and MEK/ERK dependent) versus adenosine 2A receptor-induced pMF (i.e., TrkB synthesis and PI3K/Akt dependent).
Figures
Similar articles
-
Spinal 5-HT7 receptors and protein kinase A constrain intermittent hypoxia-induced phrenic long-term facilitation.Neuroscience. 2013 Oct 10;250:632-43. doi: 10.1016/j.neuroscience.2013.06.068. Epub 2013 Jul 11. Neuroscience. 2013. PMID: 23850591 Free PMC article.
-
Systemic inflammation inhibits serotonin receptor 2-induced phrenic motor facilitation upstream from BDNF/TrkB signaling.J Neurophysiol. 2018 Jun 1;119(6):2176-2185. doi: 10.1152/jn.00378.2017. Epub 2018 Mar 7. J Neurophysiol. 2018. PMID: 29513151 Free PMC article.
-
BDNF-induced phrenic motor facilitation shifts from PKCθ to ERK dependence with mild systemic inflammation.J Neurophysiol. 2023 Feb 1;129(2):455-464. doi: 10.1152/jn.00345.2022. Epub 2023 Jan 25. J Neurophysiol. 2023. PMID: 36695529 Free PMC article.
-
Circulatory control of phrenic motor plasticity.Respir Physiol Neurobiol. 2019 Jul;265:19-23. doi: 10.1016/j.resp.2019.01.004. Epub 2019 Jan 11. Respir Physiol Neurobiol. 2019. PMID: 30639504 Free PMC article. Review.
-
NADPH oxidase activity is necessary for acute intermittent hypoxia-induced phrenic long-term facilitation.J Physiol. 2009 May 1;587(Pt 9):1931-42. doi: 10.1113/jphysiol.2008.165597. Epub 2009 Feb 23. J Physiol. 2009. PMID: 19237427 Free PMC article. Review.
Cited by
-
Differential mechanisms are required for phrenic long-term facilitation over the course of motor neuron loss following CTB-SAP intrapleural injections.Exp Neurol. 2020 Dec;334:113460. doi: 10.1016/j.expneurol.2020.113460. Epub 2020 Sep 9. Exp Neurol. 2020. PMID: 32916172 Free PMC article.
-
Systemic LPS induces spinal inflammatory gene expression and impairs phrenic long-term facilitation following acute intermittent hypoxia.J Appl Physiol (1985). 2013 Apr;114(7):879-87. doi: 10.1152/japplphysiol.01347.2012. Epub 2013 Jan 17. J Appl Physiol (1985). 2013. PMID: 23329821 Free PMC article.
-
Spinal 5-HT7 receptors induce phrenic motor facilitation via EPAC-mTORC1 signaling.J Neurophysiol. 2015 Sep;114(3):2015-22. doi: 10.1152/jn.00374.2015. Epub 2015 Aug 12. J Neurophysiol. 2015. PMID: 26269554 Free PMC article.
-
Cervical spinal 5-HT2A and 5-HT2B receptors are both necessary for moderate acute intermittent hypoxia-induced phrenic long-term facilitation.J Appl Physiol (1985). 2019 Aug 1;127(2):432-443. doi: 10.1152/japplphysiol.01113.2018. Epub 2019 Jun 20. J Appl Physiol (1985). 2019. PMID: 31219768 Free PMC article.
-
Increased spinal adenosine impairs phrenic long-term facilitation in aging rats.J Appl Physiol (1985). 2023 Jun 1;134(6):1537-1548. doi: 10.1152/japplphysiol.00197.2023. Epub 2023 May 11. J Appl Physiol (1985). 2023. PMID: 37167263 Free PMC article.
References
-
- Bach KB, Mitchell GS. Hypoxia-induced long-term facilitation of respiratory activity is serotonin dependent. Respir Physiol 104: 251–260, 1996 - PubMed
-
- Baker-Herman TL, Fuller DD, Bavis RW, Zabka AG, Golder FJ, Doperalski NJ, Johnson RA, Watters JJ, Mitchell GS. BDNF is necessary and sufficient for spinal respiratory plasticity following intermittent hypoxia. Nat Neurosci 7: 48, 2004 - PubMed
-
- Behan M, Zabka AG, Mitchell GS. Age and gender effects on serotonin-dependent plasticity in respiratory motor control. Respir Physiol Neurobiol 131: 65–77, 2002 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous