

SurA is involved in the targeting to the outer membrane of a Tat signal sequence-anchored protein
- PMID: 22961852
- PMCID: PMC3486411
- DOI: 10.1128/JB.01419-12
SurA is involved in the targeting to the outer membrane of a Tat signal sequence-anchored protein
Abstract
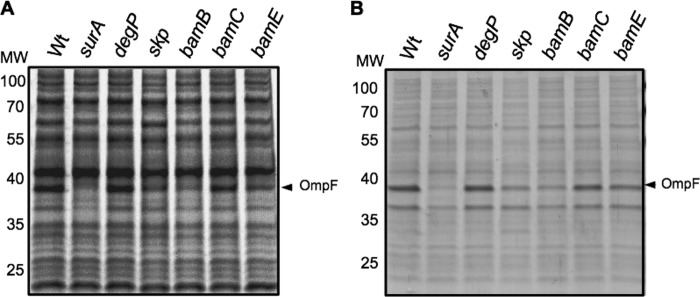
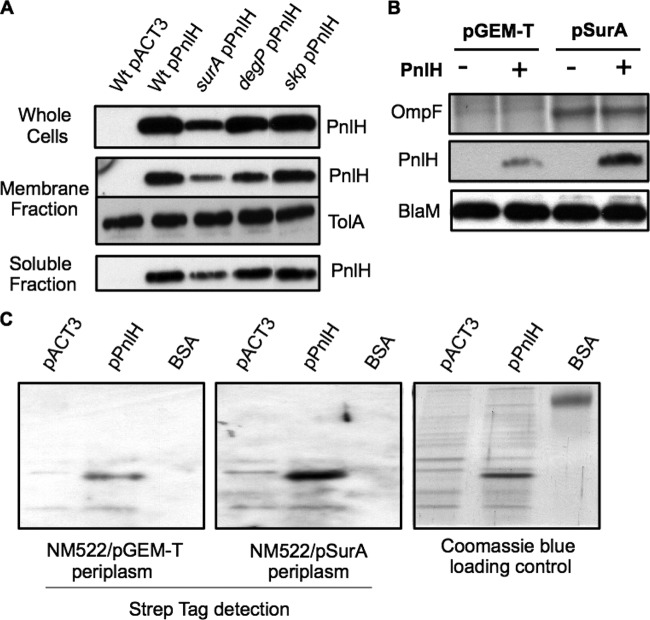
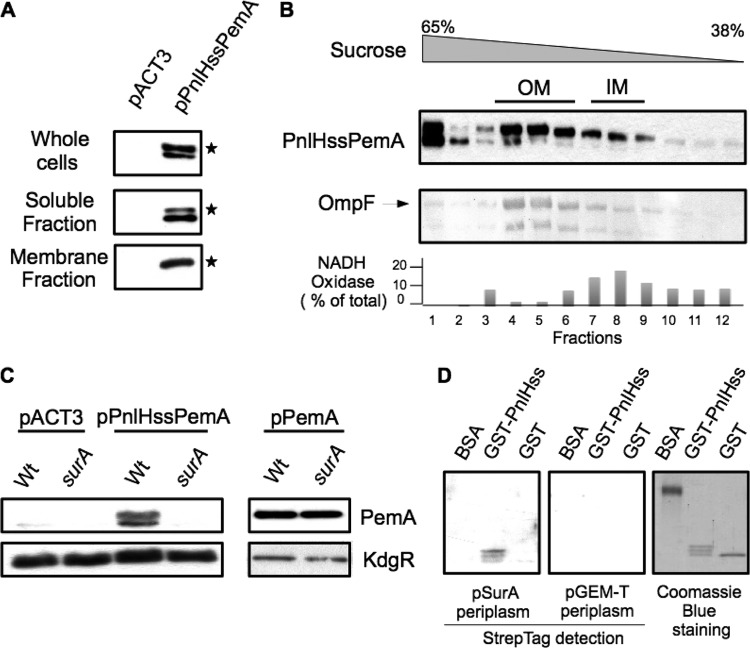
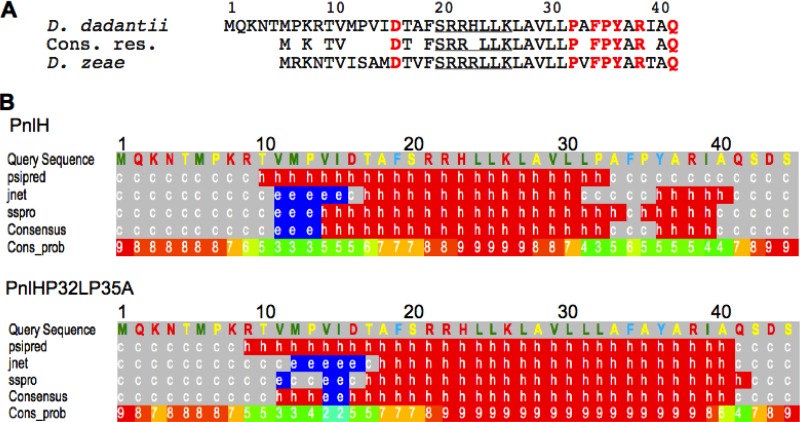
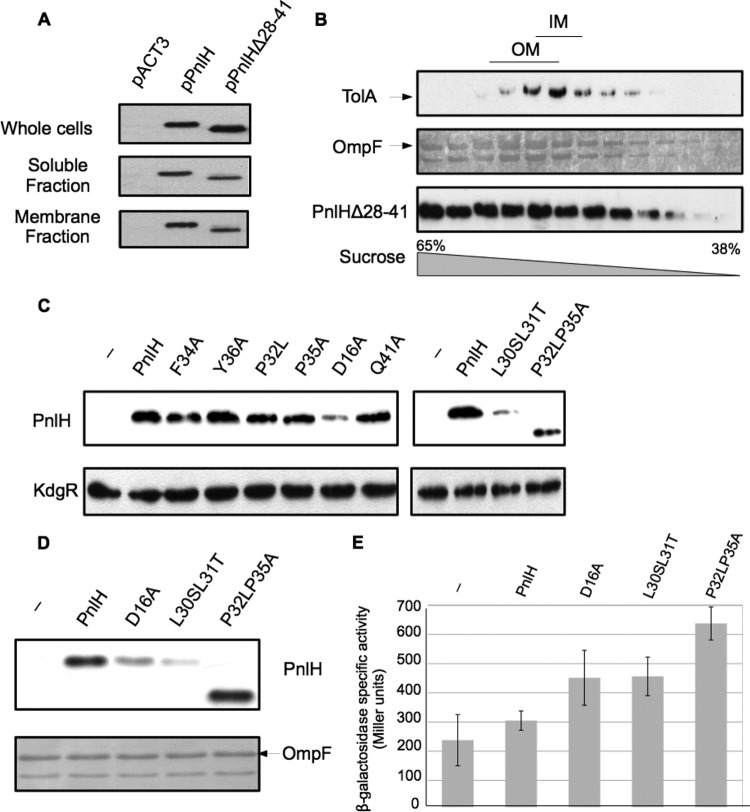
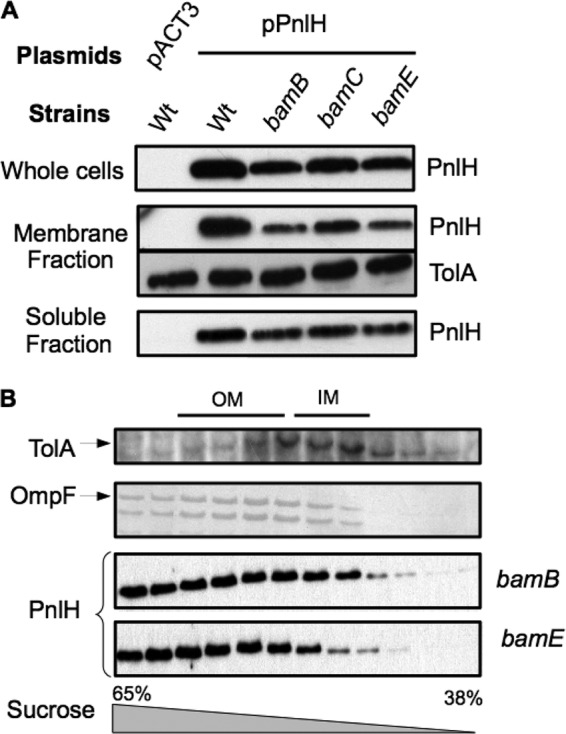
The twin arginine translocation (Tat) pathway exports folded proteins from the cytoplasm to the periplasm of bacteria. The targeting of the exported proteins to the Tat pathway relies on a specific amino-terminal signal sequence, which is cleaved after exportation. In the phytopathogen Dickeya dadantii, the pectin lyase homologue PnlH is exported by the Tat pathway without cleavage of its signal sequence, which anchors PnlH into the outer membrane. In proteobacteria, the vast majority of outer membrane proteins consists of β-barrel proteins and lipoproteins. Thus, PnlH represents a new kind of outer membrane protein. In Escherichia coli, periplasmic chaperones SurA, Skp, and DegP work together with the β-barrel assembly machinery (Bam) to target and insert β-barrel proteins into the outer membrane. In this work, we showed that SurA is required for an efficient targeting of PnlH to the outer membrane. Moreover, we were able to detect an in vitro interaction between SurA and the PnlH signal sequence. Since the PnlH signal sequence contains a highly hydrophobic region, we propose that SurA protects it from the hydrophobic periplasm during targeting of PnlH to the outer membrane. We also studied the nature of the information carried by the PnlH signal sequence responsible for its targeting to the outer membrane after exportation by the Tat system.
Figures
Similar articles
-
Bacterial Outer Membrane Proteins Are Targeted to the Bam Complex by Two Parallel Mechanisms.mBio. 2021 May 4;12(3):e00597-21. doi: 10.1128/mBio.00597-21. mBio. 2021. PMID: 33947759 Free PMC article.
-
The fimbrial usher FimD follows the SurA-BamB pathway for its assembly in the outer membrane of Escherichia coli.J Bacteriol. 2011 Oct;193(19):5222-30. doi: 10.1128/JB.05585-11. Epub 2011 Jul 22. J Bacteriol. 2011. PMID: 21784935 Free PMC article.
-
Novel mechanism of outer membrane targeting of proteins in Gram-negative bacteria.Mol Microbiol. 2008 Sep;69(6):1349-57. doi: 10.1111/j.1365-2958.2008.06366.x. Epub 2008 Jul 11. Mol Microbiol. 2008. PMID: 18643934
-
The role of SurA factor in outer membrane protein transport and virulence.Int J Med Microbiol. 2010 Nov;300(7):421-8. doi: 10.1016/j.ijmm.2010.04.012. Epub 2010 May 6. Int J Med Microbiol. 2010. PMID: 20447864 Review.
-
Periplasmic quality control in biogenesis of outer membrane proteins.Biochem Soc Trans. 2015 Apr;43(2):133-8. doi: 10.1042/BST20140217. Biochem Soc Trans. 2015. PMID: 25849907 Review.
Cited by
-
A microbial sensor for discovering structural probes of protein misfolding and aggregation.Prion. 2013 Mar-Apr;7(2):151-6. doi: 10.4161/pri.23328. Epub 2013 Jan 28. Prion. 2013. PMID: 23357829 Free PMC article. Review.
-
Microbial peptidyl-prolyl cis/trans isomerases (PPIases): virulence factors and potential alternative drug targets.Microbiol Mol Biol Rev. 2014 Sep;78(3):544-71. doi: 10.1128/MMBR.00015-14. Microbiol Mol Biol Rev. 2014. PMID: 25184565 Free PMC article. Review.
-
An engineered genetic selection for ternary protein complexes inspired by a natural three-component hitchhiker mechanism.Sci Rep. 2014 Dec 22;4:7570. doi: 10.1038/srep07570. Sci Rep. 2014. PMID: 25531212 Free PMC article.
References
-
- Alcock FH, et al. 2008. Conserved substrate binding by chaperones in the bacterial periplasm and the mitochondrial intermembrane space. Biochem. J. 409:377–387 - PubMed
-
- Allen JWA, Tomlinson EJ, Hong L, Ferguson SJ. 2002. The Escherichia coli cytochrome c maturation (Ccm) system does not detectably attach heme to single cysteine variants of an apocytochrome c. J. Biol. Chem. 277:33559–33563 - PubMed
-
- de Cock H, Struyvé M, Kleerebezem M, van der Krift T, Tommassen J. 1997. Role of the carboxy-terminal phenylalanine in the biogenesis of outer membrane protein PhoE of Escherichia coli K-12. J. Mol. Biol. 269:473–478 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
