

Hyperosmotic stress response of Campylobacter jejuni
- PMID: 22961853
- PMCID: PMC3486418
- DOI: 10.1128/JB.01409-12
Hyperosmotic stress response of Campylobacter jejuni
Abstract
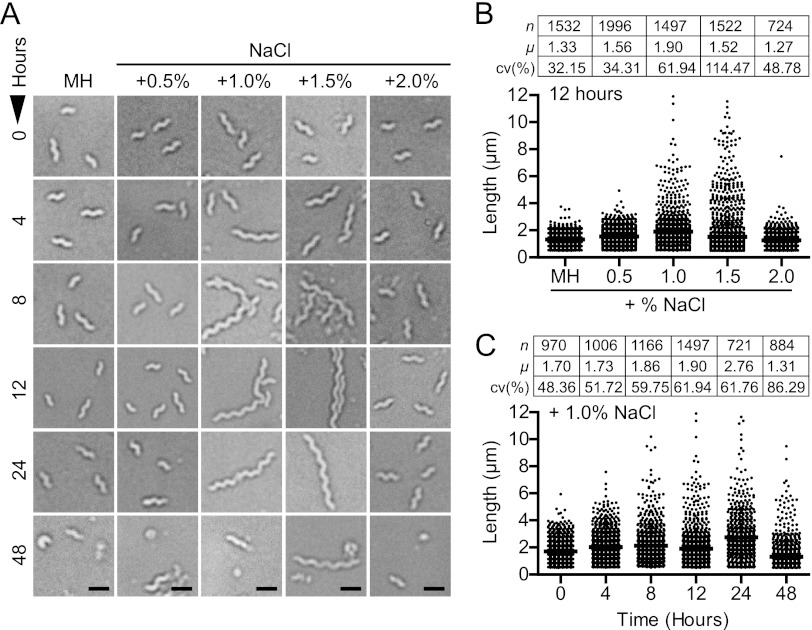
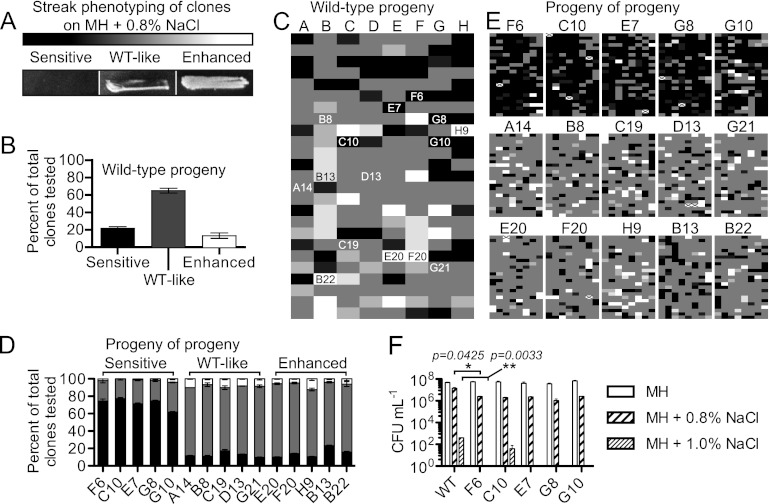
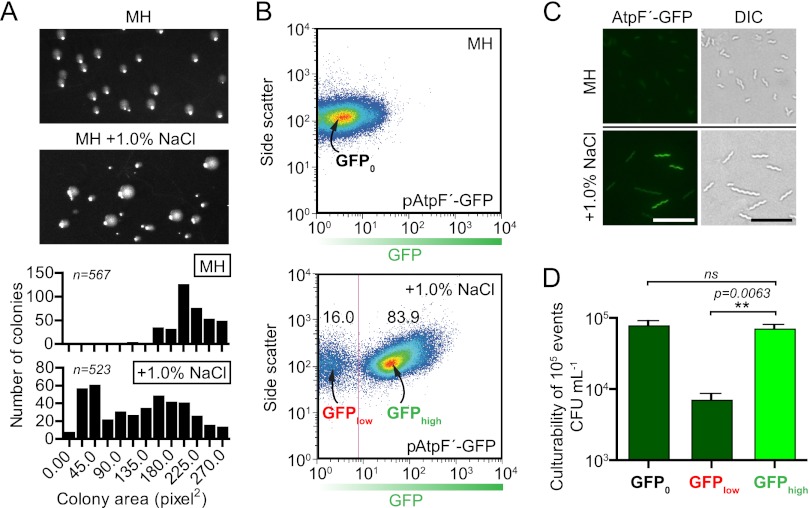
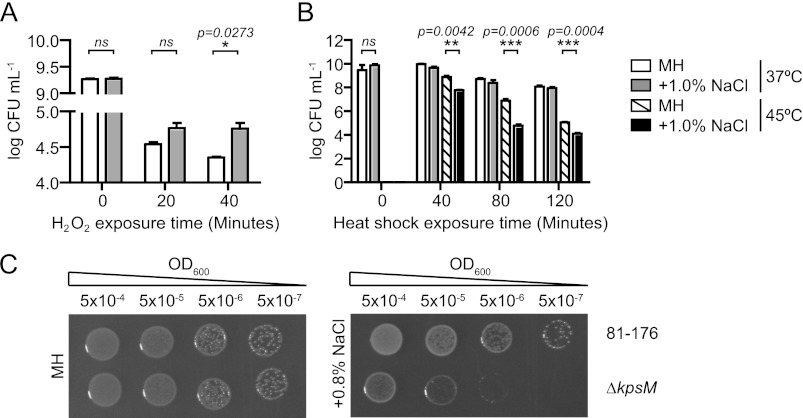
The diarrheal pathogen Campylobacter jejuni and other gastrointestinal bacteria encounter changes in osmolarity in the environment, through exposure to food processing, and upon entering host organisms, where osmotic adaptation can be associated with virulence. In this study, growth profiles, transcriptomics, and phenotypic, mutant, and single-cell analyses were used to explore the effects of hyperosmotic stress exposure on C. jejuni. Increased growth inhibition correlated with increased osmotic concentration, with both ionic and nonionic stressors inhibiting growth at 0.620 total osmol liter(-1). C. jejuni adaptation to a range of osmotic stressors and concentrations was accompanied by severe filamentation in subpopulations, with microscopy indicating septum formation and phenotypic diversity between individual cells in a filament. Population heterogeneity was also exemplified by the bifurcation of colony morphology into small and large variants on salt stress plates. Flow cytometry of C. jejuni harboring green fluorescent protein (GFP) fused to the ATP synthase promoter likewise revealed bimodal subpopulations under hyperosmotic stress. We also identified frequent hyperosmotic stress-sensitive variants within the clonal wild-type population propagated on standard laboratory medium. Microarray analysis following hyperosmotic upshift revealed enhanced expression of heat shock genes and genes encoding enzymes for synthesis of potential osmoprotectants and cross-protective induction of oxidative stress genes. The capsule export gene kpsM was also upregulated, and an acapsular mutant was defective for growth under hyperosmotic stress. For C. jejuni, an organism lacking most conventional osmotic response factors, these data suggest an unusual hyperosmotic stress response, including likely "bet-hedging" survival strategies relying on the presence of stress-fit individuals in a heterogeneous population.
Figures



References
-
- Andersen MT, et al. 2005. Diverse roles for HspR in Campylobacter jejuni revealed by the proteome, transcriptome and phenotypic characterization of an hspR mutant. Microbiology 151:905–915 - PubMed
-
- Bacon DJ, et al. 2001. A phase-variable capsule is involved in virulence of Campylobacter jejuni 81-176. Mol. Microbiol. 40:769–777 - PubMed
-
- Bonis M, Ecobichon C, Guadagnini S, Prevost MC, Boneca IG. 2010. A M23B family metallopeptidase of Helicobacter pylori required for cell shape, pole formation and virulence. Mol. Microbiol. 78:809–819 - PubMed
-
- Boulos L, Prevost M, Barbeau B, Coallier J, Desjardins R. 1999. LIVE/DEAD BacLight: application of a new rapid staining method for direct enumeration of viable and total bacteria in drinking water. J. Microbiol. Methods 37:77–86 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
