

Induction of cancerous stem cells during embryonic stem cell differentiation
- PMID: 22961983
- PMCID: PMC3481281
- DOI: 10.1074/jbc.M112.372557
Induction of cancerous stem cells during embryonic stem cell differentiation
Abstract
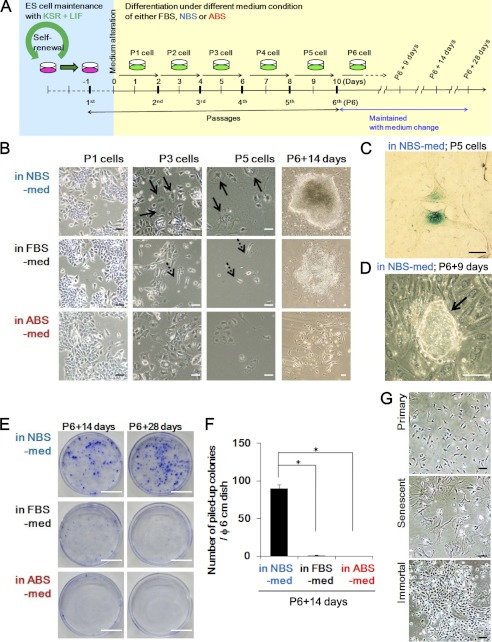
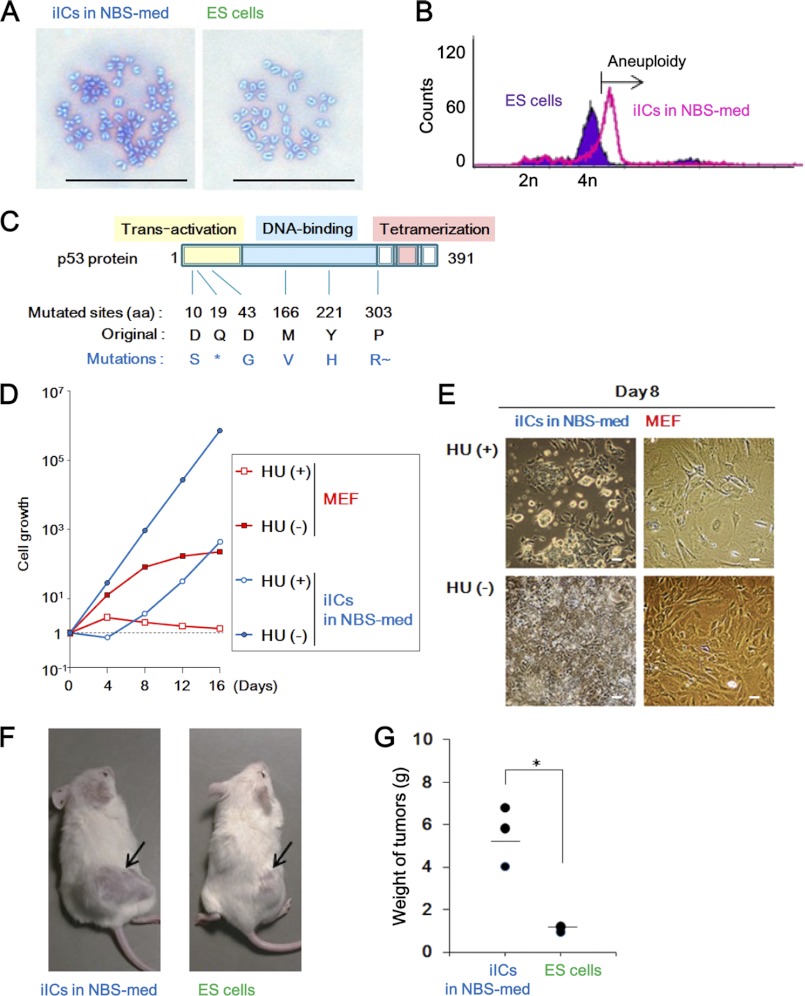
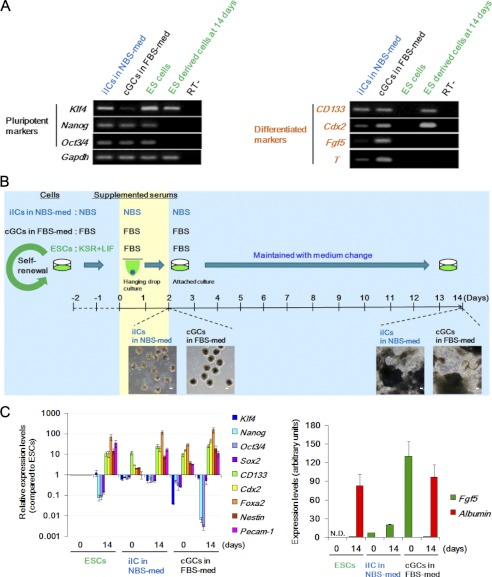
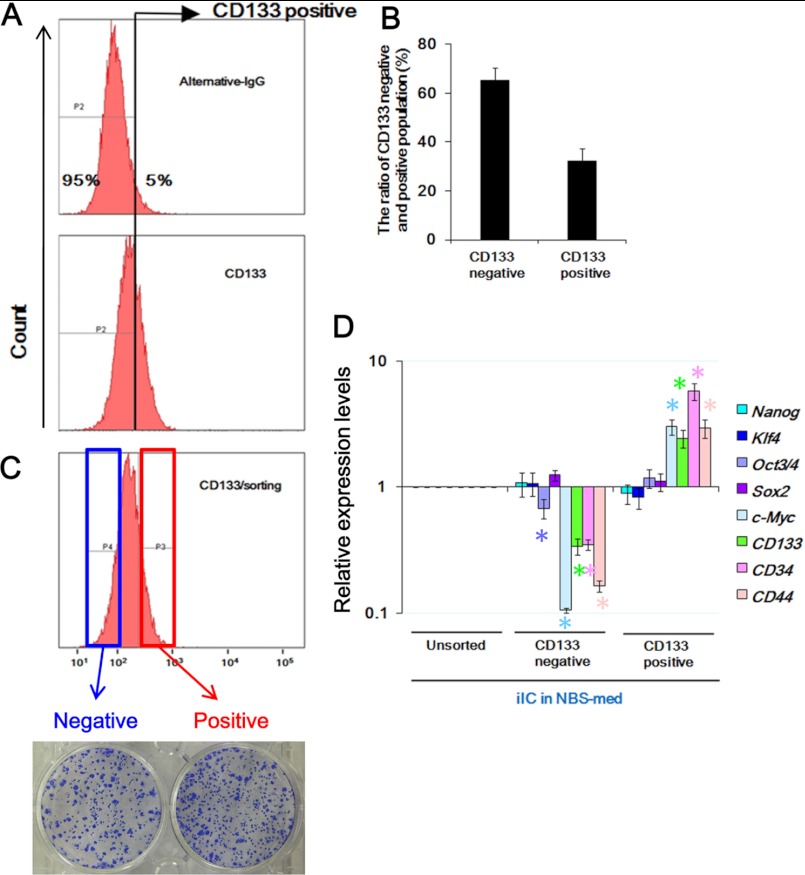
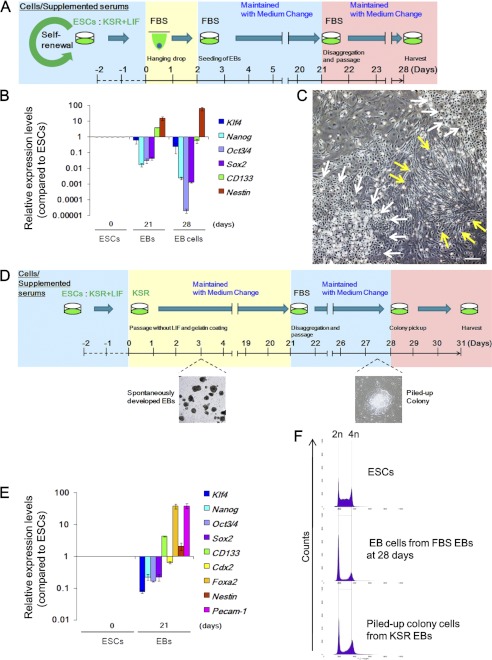
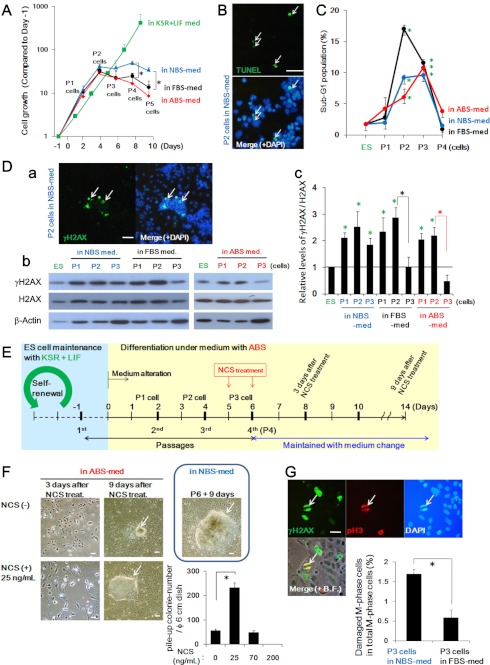
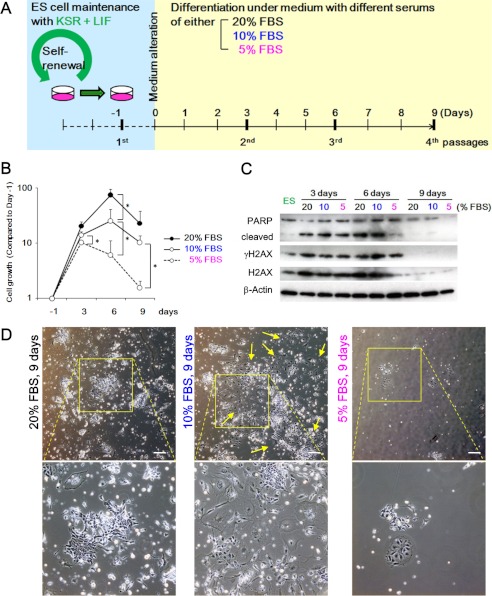
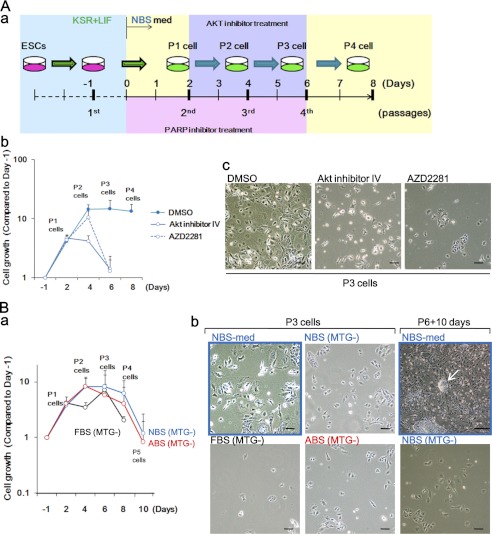
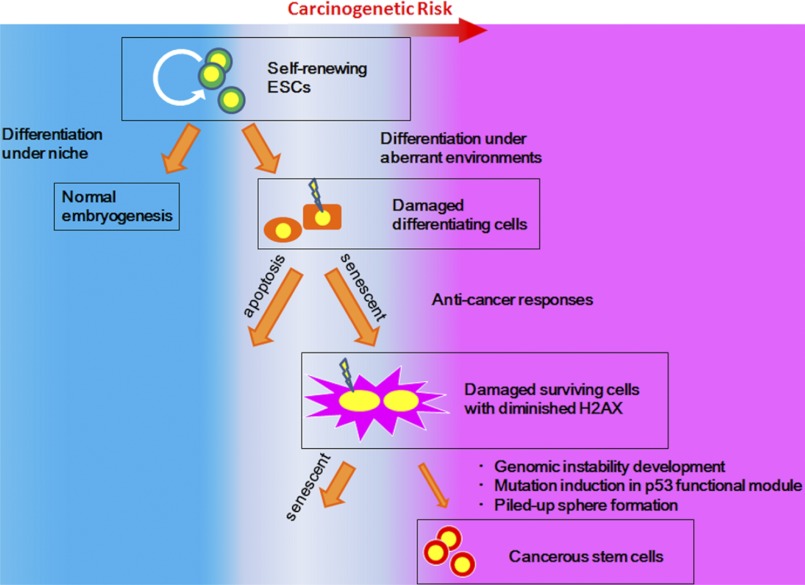
Stem cell maintenance depends on their surrounding microenvironment, and aberrancies in the environment have been associated with tumorigenesis. However, it remains to be elucidated whether an environmental aberrancy can act as a carcinogenic stress for cellular transformation of differentiating stem cells into cancer stem cells. Here, utilizing mouse embryonic stem cells as a model, it was illustrated that environmental aberrancy during differentiation leads to the emergence of pluripotent cells showing cancerous characteristics. Analogous to precancerous stages, DNA lesions were spontaneously accumulated during embryonic stem cell differentiation under aberrational environments, which activates barrier responses such as senescence and apoptosis. However, overwhelming such barrier responses, piled-up spheres were subsequently induced from the previously senescent cells. The sphere cells exhibit aneuploidy and dysfunction of the Arf-p53 module as well as enhanced tumorigenicity and a strong self-renewal capacity, suggesting development of cancerous stem cells. Our current study suggests that stem cells differentiating in an aberrational environment are at risk of cellular transformation into malignant counterparts.
Figures
Similar articles
-
Cultivation and identification of colon cancer stem cell-derived spheres from the Colo205 cell line.Braz J Med Biol Res. 2012 Mar;45(3):197-204. doi: 10.1590/s0100-879x2012007500015. Epub 2012 Feb 9. Braz J Med Biol Res. 2012. PMID: 22310487 Free PMC article.
-
CD133(+) gallbladder carcinoma cells exhibit self-renewal ability and tumorigenicity.World J Gastroenterol. 2011 Jun 28;17(24):2965-71. doi: 10.3748/wjg.v17.i24.2965. World J Gastroenterol. 2011. PMID: 21734809 Free PMC article.
-
Nodal/Activin signaling drives self-renewal and tumorigenicity of pancreatic cancer stem cells and provides a target for combined drug therapy.Cell Stem Cell. 2011 Nov 4;9(5):433-46. doi: 10.1016/j.stem.2011.10.001. Cell Stem Cell. 2011. PMID: 22056140
-
[Updates on study of glioma stem cells].Zhonghua Bing Li Xue Za Zhi. 2007 Mar;36(3):201-3. Zhonghua Bing Li Xue Za Zhi. 2007. PMID: 17535692 Review. Chinese. No abstract available.
-
CD133 as a marker for regulation and potential for targeted therapies in glioblastoma multiforme.Neurosurg Clin N Am. 2012 Jul;23(3):391-405. doi: 10.1016/j.nec.2012.04.011. Epub 2012 Jun 5. Neurosurg Clin N Am. 2012. PMID: 22748652 Review.
Cited by
-
Integrins in cancer stem cells.Front Cell Dev Biol. 2024 Aug 21;12:1434378. doi: 10.3389/fcell.2024.1434378. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39239559 Free PMC article. Review.
-
Unraveling the journey of cancer stem cells from origin to metastasis.Biochim Biophys Acta Rev Cancer. 2019 Jan;1871(1):50-63. doi: 10.1016/j.bbcan.2018.10.006. Epub 2018 Nov 9. Biochim Biophys Acta Rev Cancer. 2019. PMID: 30419314 Free PMC article. Review.
-
Targeting CREB-binding protein (CBP) abrogates colorectal cancer stemness through epigenetic regulation of C-MYC.Cancer Gene Ther. 2024 Nov;31(11):1734-1748. doi: 10.1038/s41417-024-00838-9. Epub 2024 Oct 2. Cancer Gene Ther. 2024. PMID: 39358564
-
CD44 standard and CD44v10 isoform expression on leukemia cells distinctly influences niche embedding of hematopoietic stem cells.J Hematol Oncol. 2014 Mar 31;7:29. doi: 10.1186/1756-8722-7-29. J Hematol Oncol. 2014. PMID: 24684724 Free PMC article.
-
A comprehensive analysis of radiosensitization targets; functional inhibition of DNA methyltransferase 3B radiosensitizes by disrupting DNA damage regulation.Sci Rep. 2015 Dec 15;5:18231. doi: 10.1038/srep18231. Sci Rep. 2015. PMID: 26667181 Free PMC article.
References
-
- Wicha M. S., Liu S., Dontu G. (2006) Cancer stem cells. An old idea, a paradigm shift. Cancer Res. 66, 1883–1890 - PubMed
-
- Werbowetski-Ogilvie T. E., Bhatia M. (2008) Pluripotent human stem cell lines. What we can learn about cancer initiation. Trends Mol. Med. 14, 323–332 - PubMed
-
- Chiba T., Zheng Y. W., Kita K., Yokosuka O., Saisho H., Onodera M., Miyoshi H., Nakano M., Zen Y., Nakanuma Y., Nakauchi H., Iwama A., Taniguchi H. (2007) Enhanced self-renewal capability in hepatic stem/progenitor cells drives cancer initiation. Gastroenterology 133, 937–950 - PubMed
-
- Bartkova J., Horejsí Z., Koed K., Krämer A., Tort F., Zieger K., Guldberg P., Sehested M., Nesland J. M., Lukas C., Ørntoft T., Lukas J., Bartek J. (2005) DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature 434, 864–870 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
