

Molecular basis for glucocorticoid induction of the Kruppel-like factor 9 gene in hippocampal neurons
- PMID: 22962255
- PMCID: PMC3473204
- DOI: 10.1210/en.2012-1303
Molecular basis for glucocorticoid induction of the Kruppel-like factor 9 gene in hippocampal neurons
Abstract
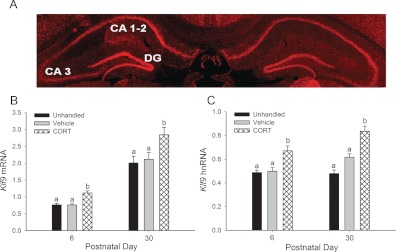
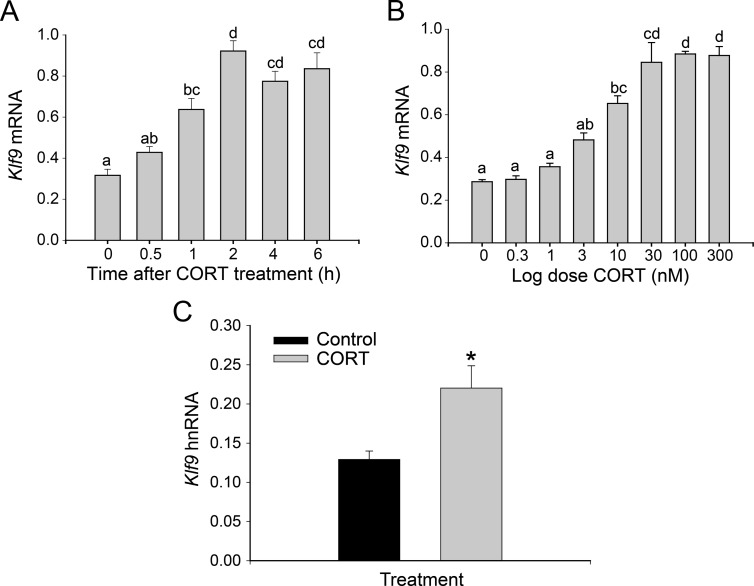
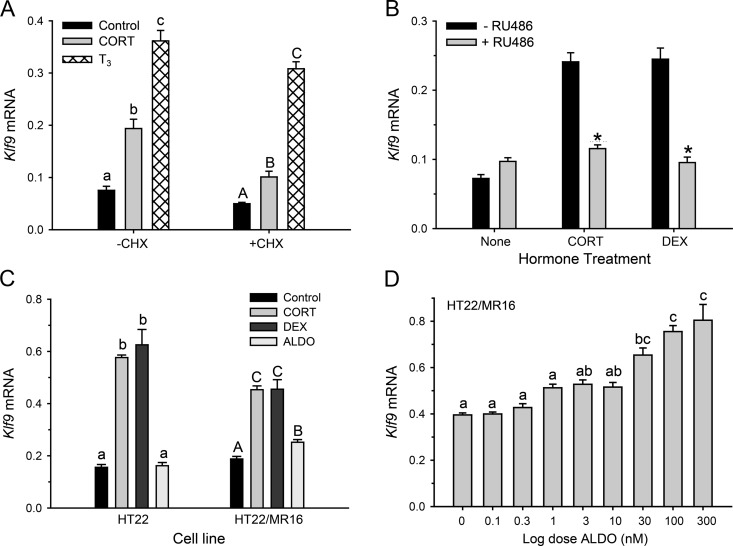
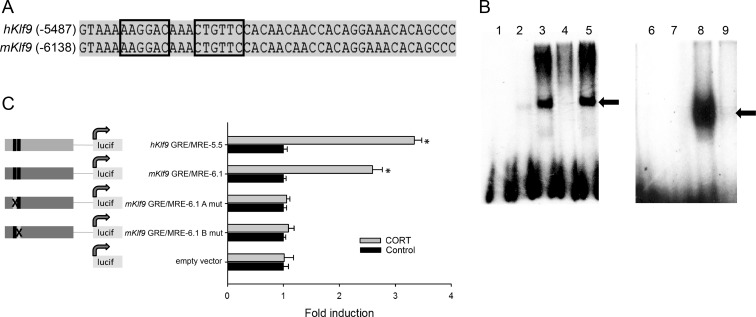
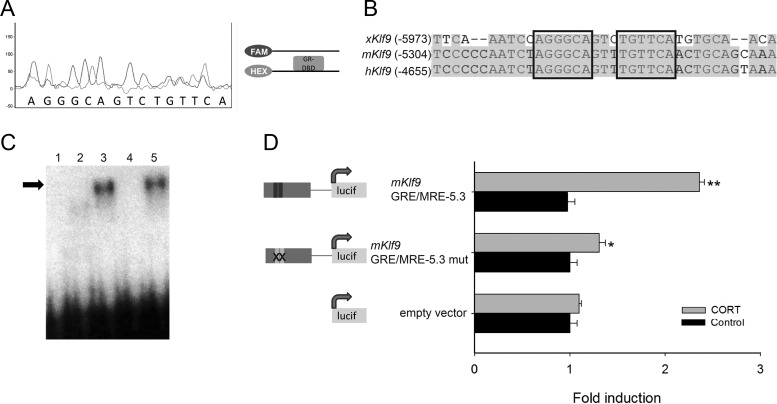
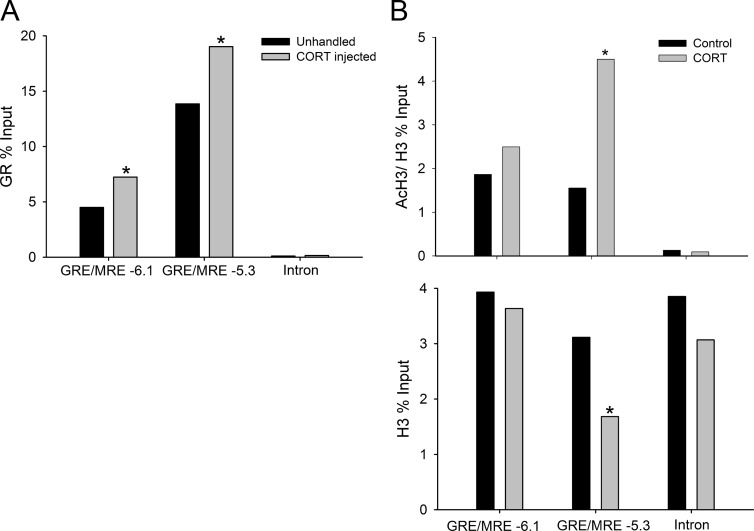
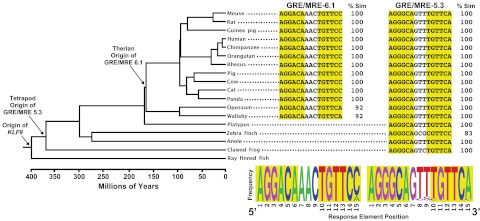
Stress has complex effects on hippocampal structure and function, which consequently affects learning and memory. These effects are mediated in part by circulating glucocorticoids (GC) acting via the intracellular GC receptor (GR) and mineralocorticoid receptor (MR). Here, we investigated GC regulation of Krüppel-like factor 9 (KLF9), a transcription factor implicated in neuronal development and plasticity. Injection of corticosterone (CORT) in postnatal d 6 and 30 mice increased Klf9 mRNA and heteronuclear RNA by 1 h in the hippocampal region. Treatment of the mouse hippocampal cell line HT-22 with CORT caused a time- and dose-dependent increase in Klf9 mRNA. The CORT induction of Klf9 was resistant to protein synthesis inhibition, suggesting that Klf9 is a direct CORT-response gene. In support of this hypothesis, we identified two GR/MR response elements (GRE/MRE) located -6.1 and -5.3 kb relative to the transcription start site, and we verified their functionality by enhancer-reporter, gel shift, and chromatin immunoprecipitation assays. The -5.3-kb GRE/MRE is largely conserved across tetrapods, but conserved orthologs of the -6.1-kb GRE/MRE were only detected in therian mammals. GC treatment caused recruitment of the GR, histone hyperacetylation, and nucleosome removal at Klf9 upstream regions. Our findings support a predominant role for GR, with a minor contribution of MR, in the direct regulation of Klf9 acting via two GRE/MRE located in the 5'-flanking region of the gene. KLF9 may play a key role in GC actions on hippocampal development and plasticity.
Figures
References
-
- Joëls M, Pu Z, Wiegert O, Oitzl MS, Krugers HJ. 2006. Learning under stress: how does it work? Trends Cogn Sci 10:152–158 - PubMed
-
- Reul JM, de Kloet ER. 1985. Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. Endocrinology 117:2505–2511 - PubMed
-
- De Kloet ER, Vreugdenhil E, Oitzl MS, Joëls M. 1998. Brain corticosteroid receptor balance in health and disease. Endocr Rev 19:269–301 - PubMed
-
- Puymirat J. 1992. Thyroid receptors in the rat brain. Prog Neurobiol 39:281–294 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
