

CIB1 prevents nuclear GAPDH accumulation and non-apoptotic tumor cell death via AKT and ERK signaling
- PMID: 22964641
- PMCID: PMC3530648
- DOI: 10.1038/onc.2012.408
CIB1 prevents nuclear GAPDH accumulation and non-apoptotic tumor cell death via AKT and ERK signaling
Abstract
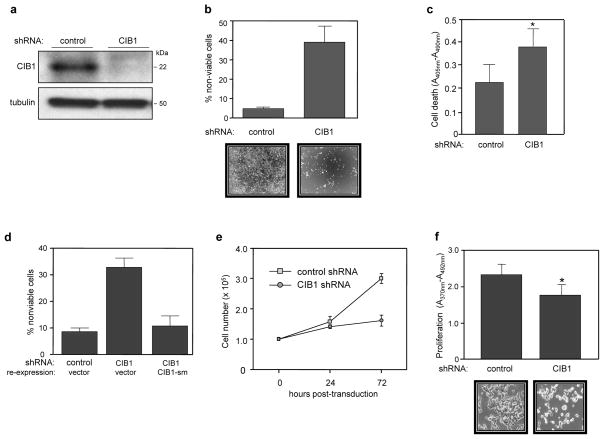
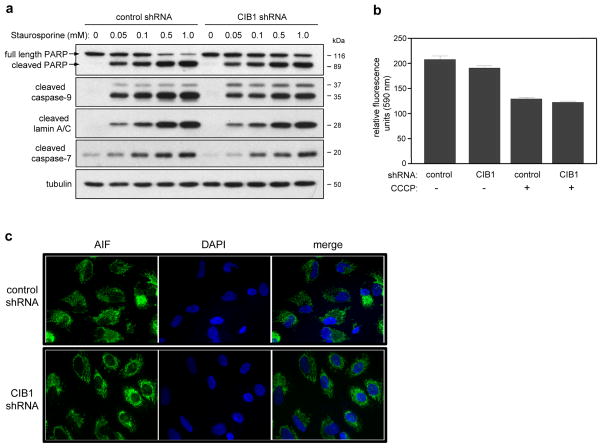
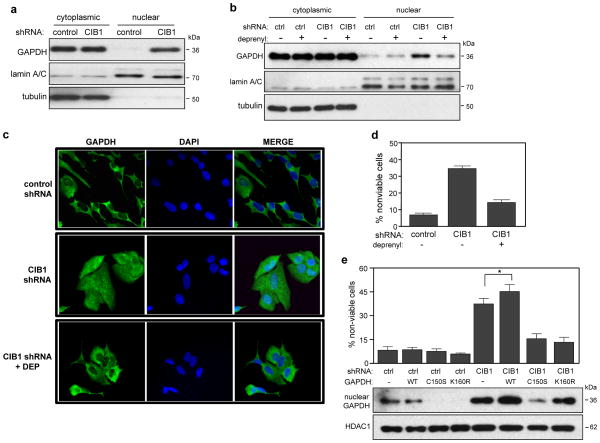
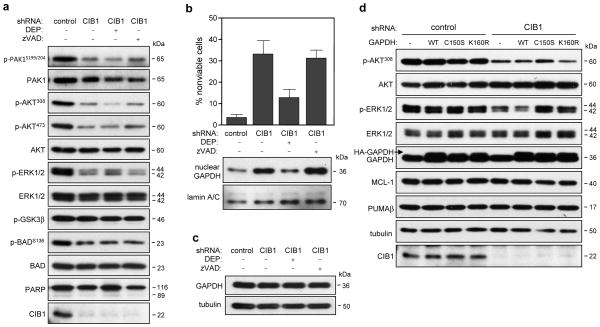
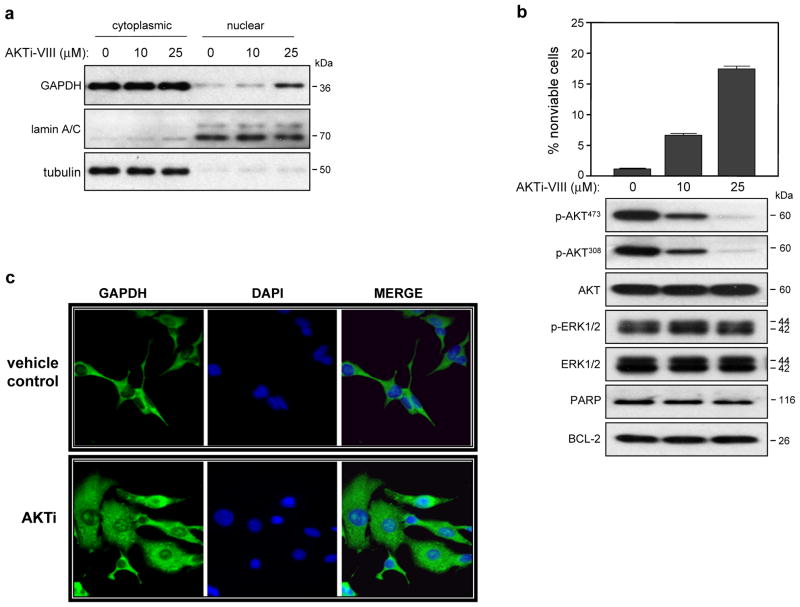
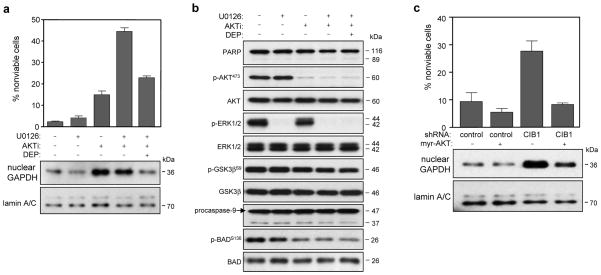
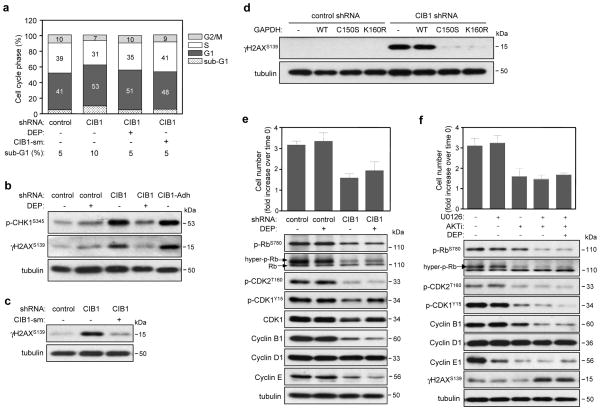
CIB1 is a 22-kDa regulatory protein previously implicated in cell survival and proliferation. However, the mechanism by which CIB1 regulates these processes is poorly defined. Here, we report that CIB1 depletion in SK-N-SH neuroblastoma and MDA-MB-468 breast cancer cells promotes non-apoptotic, caspase-independent cell death that is not initiated by increased outer mitochondrial membrane permeability or translocation of apoptosis-inducing factor to the nucleus. Instead, cell death requires nuclear GAPDH accumulation. Furthermore, CIB1 depletion disrupts two commonly dysregulated, oncogenic pathways-PI3K/AKT and Ras/MEK/ERK, resulting in a synergistic mechanism of cell death, which was mimicked by simultaneous pharmacological inhibition of both pathways, but not either pathway alone. In defining each pathway's contributions, we found that AKT inhibition alone maximally induced GAPDH nuclear accumulation, whereas MEK/ERK inhibition alone had no effect on GAPDH localization. Concurrent GAPDH nuclear accumulation and ERK inhibition were required, however, to induce a significant DNA damage response, which was critical to subsequent cell death. Collectively, our results indicate that CIB1 is uniquely positioned to regulate PI3K/AKT and MEK/ERK signaling and that simultaneous disruption of these pathways synergistically induces a nuclear GAPDH-dependent cell death. The mechanistic insights into cell death induced by CIB1 interference suggest novel molecular targets for cancer therapy.
Figures
References
-
- McCubrey JA, Steelman LS, Abrams SL, Lee JT, Chang F, Bertrand FE, et al. Roles of the RAF/MEK/ERK and PI3K/PTEN/AKT pathways in malignant transformation and drug resistance. Adv Enzyme Regul. 2006;46:249–79. - PubMed
-
- Huynh N, Liu KH, Baldwin GS, He H. P21-activated kinase 1 stimulates colon cancer cell growth and migration/invasion via ERK- and AKT-dependent pathways. Biochim Biophys Acta. 2010;1803(9):1106–13. - PubMed
-
- Datta SR, Dudek H, Tao X, Masters S, Fu H, Gotoh Y, et al. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell. 1997;91(2):231–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
