

Increased adipose tissue hypoxia and capacity for angiogenesis and inflammation in young diet-sensitive C57 mice compared with diet-resistant FVB mice
- PMID: 22964790
- PMCID: PMC3525796
- DOI: 10.1038/ijo.2012.141
Increased adipose tissue hypoxia and capacity for angiogenesis and inflammation in young diet-sensitive C57 mice compared with diet-resistant FVB mice
Abstract
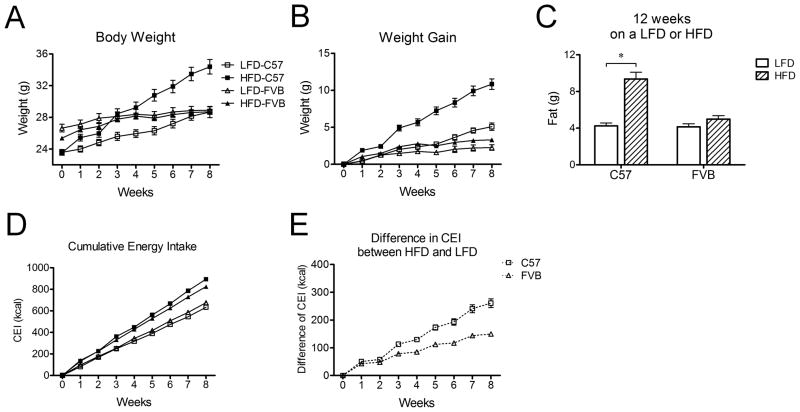
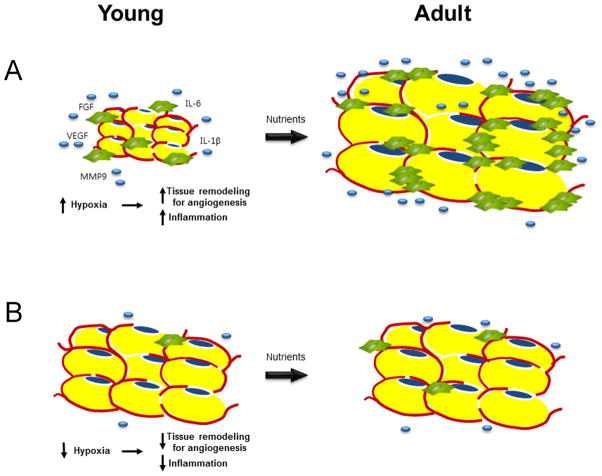
Objective: High-fat diets (HFDs) result in increased body weight. However, this is not uniform and determining the factors that make some animals or individual more susceptible to this diet-induced weight gain is a critical research question. The expansion of white adipose tissue (WAT) associated with weight gain requires high rates of angiogenesis to support the expanding tissue mass. We hypothesized that diet-induced obese (DIO) mice have a greater capacity for WAT angiogenesis and remodeling than diet-resistant (DR) mice at a young age, before age or DIO.
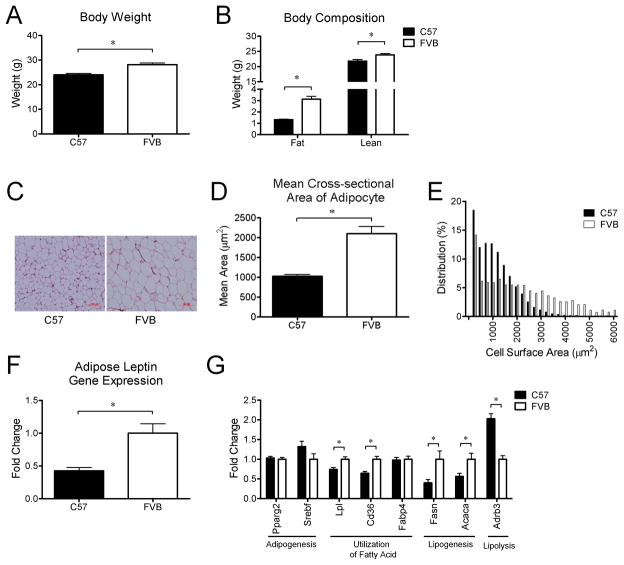
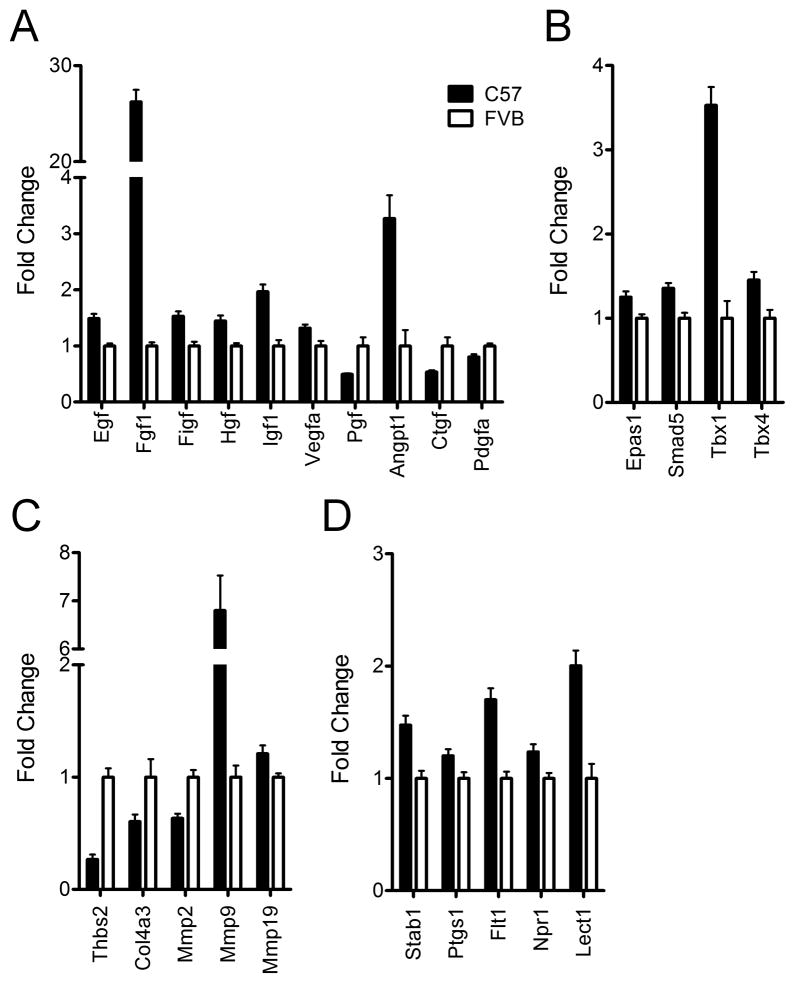
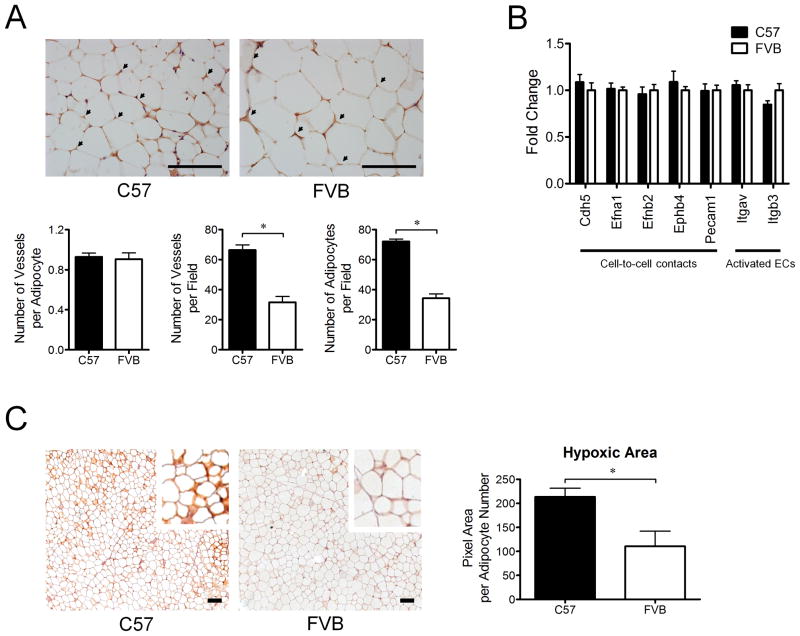
Design: We measured body weight and body composition by nuclear magnetic resonance. We compared the expression of genes related to lipid metabolism, angiogenesis and inflammation by real-time, quantitative PCR and PCR arrays. WAT morphology and distribution of adipocyte size were analyzed. The level of hypoxia and vascular density was assessed by immunohistochemistry in WAT of young mice.
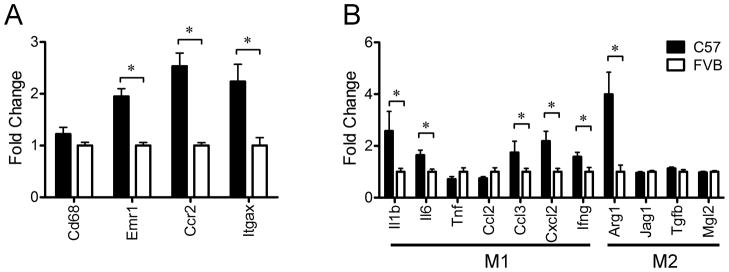
Results: C57Bl/6 mice were DIO and FVB/N (FVB) mice DR after 8 weeks on a low-fat diet or HFD. However, C57Bl/6 mice had lower body weight, lower adiposity, smaller adipocytes and decreased leptin and lipogenic genes expression in adipose tissue than FVB mice at 9 weeks of age on a chow diet. Despite having smaller adipocytes, the level of hypoxia and the expression of pro-angiogenesis genes were higher in WAT of young C57Bl/6 mice than young FVB mice. In addition, expression of genes related to macrophages and their recruitment, and to proinflammatory cytokines, was significantly higher in WAT of young C57Bl/6 mice than young FVB mice.
Conclusion: These data suggest that the potential for WAT remodeling in early period of growth is higher in C57Bl/6 mice as compared with FVB mice, and we hypothesize that it may contribute to the increased susceptibility to DIO of C57Bl/6 mice.
Conflict of interest statement
Dr. Seeley has served as a paid consultant for Novo Nordisk, Eli Lilly, Angiochem, Novartis, and Ethicon Endo-Surgery. Dr. Seeley has served as a paid speaker for Novo Nordisk, Pfizer, and Ethicon Endo-Surgery. Dr. Seeley has received research support from Novo Nordisk, Pfizer, Mannkind, Ablaris and Ethicon Endo-Surgery.
Figures
References
-
- Crandall DL, Hausman GJ, Kral JG. A review of the microcirculation of adipose tissue: anatomic, metabolic, and angiogenic perspectives. Microcirculation. 1997;4(2):211–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
