

Different cyclin types collaborate to reverse the S-phase checkpoint and permit prompt mitosis
- PMID: 22965907
- PMCID: PMC3444785
- DOI: 10.1083/jcb.201205007
Different cyclin types collaborate to reverse the S-phase checkpoint and permit prompt mitosis
Abstract
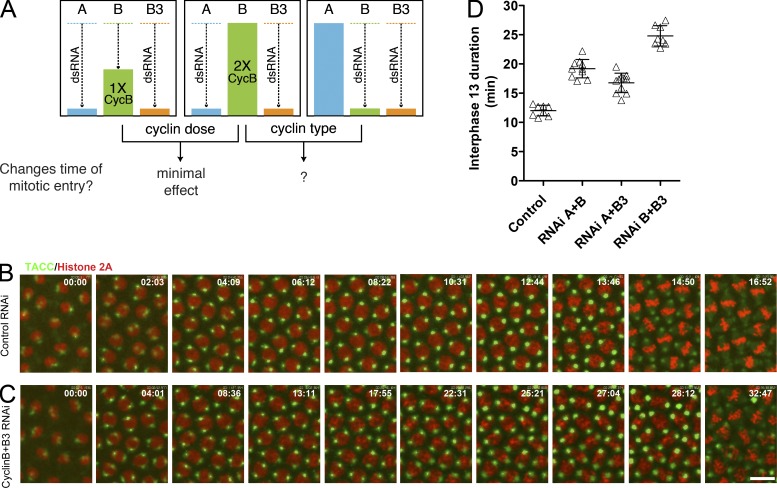
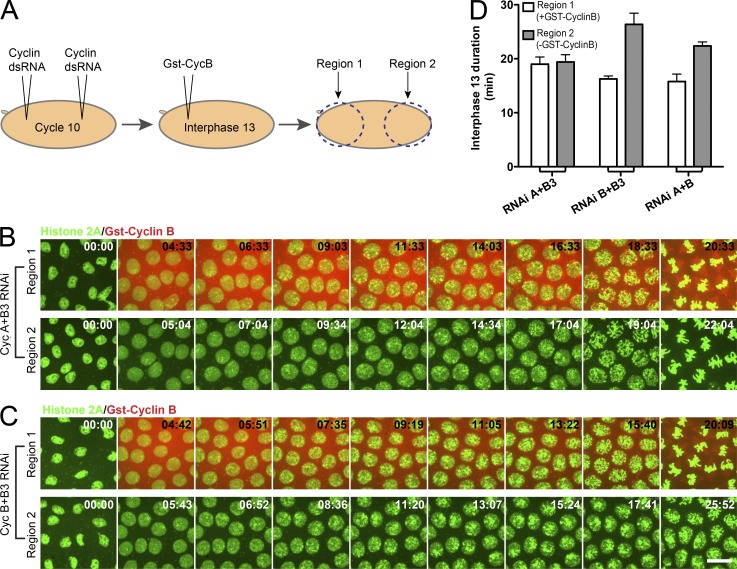
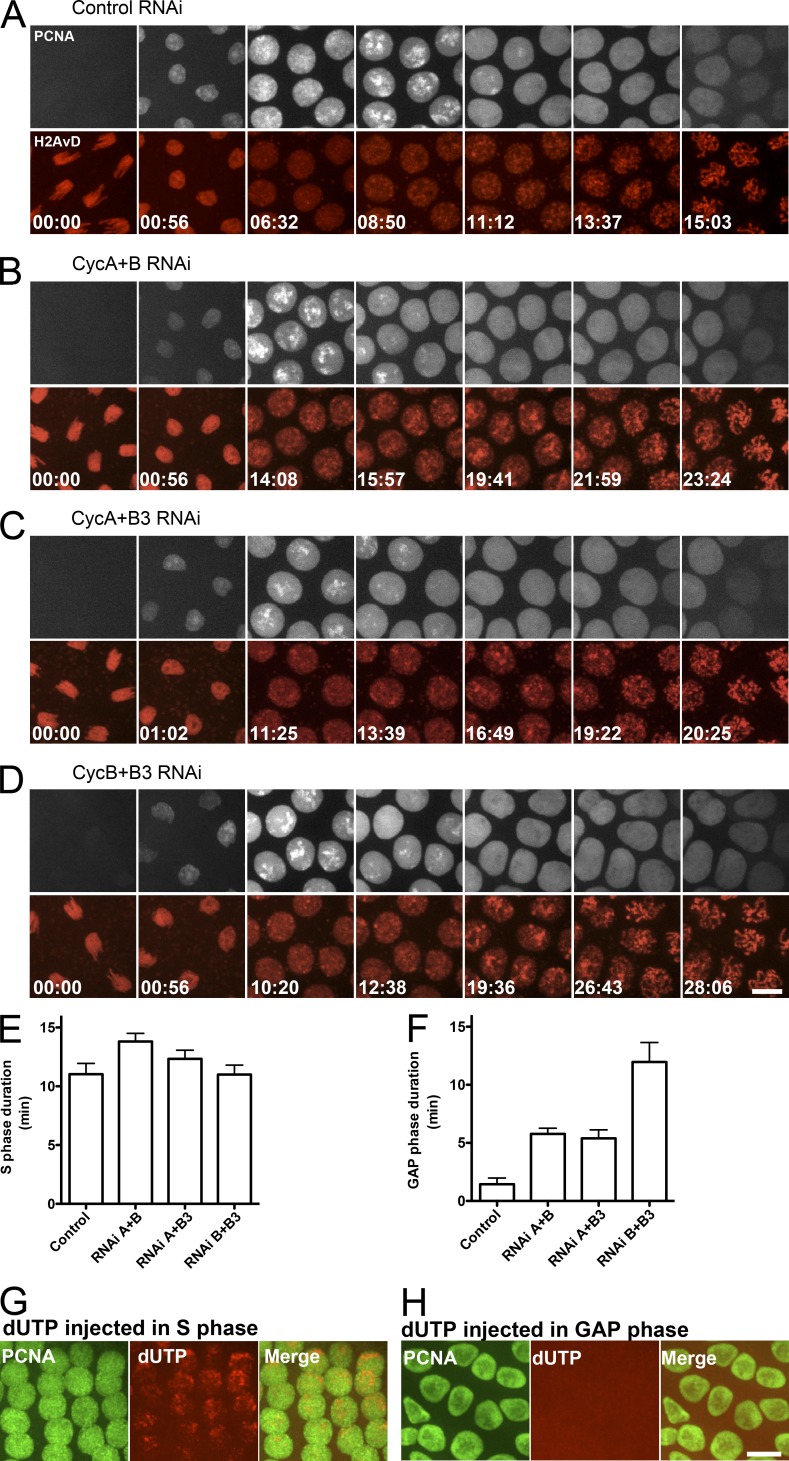
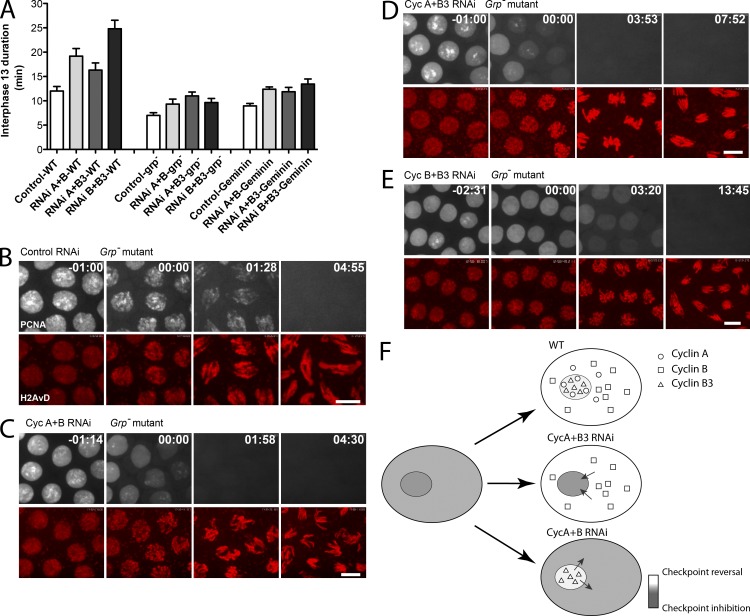
Precise timing coordinates cell proliferation with embryonic morphogenesis. As Drosophila melanogaster embryos approach cell cycle 14 and the midblastula transition, rapid embryonic cell cycles slow because S phase lengthens, which delays mitosis via the S-phase checkpoint. We probed the contributions of each of the three mitotic cyclins to this timing of interphase duration. Each pairwise RNA interference knockdown of two cyclins lengthened interphase 13 by introducing a G2 phase of a distinct duration. In contrast, pairwise cyclin knockdowns failed to introduce a G2 in embryos that lacked an S-phase checkpoint. Thus, the single remaining cyclin is sufficient to induce early mitotic entry, but reversal of the S-phase checkpoint is compromised by pairwise cyclin knockdown. Manipulating cyclin levels revealed that the diversity of cyclin types rather than cyclin level influenced checkpoint reversal. We conclude that different cyclin types have distinct abilities to reverse the checkpoint but that they collaborate to do so rapidly.
Figures
Similar articles
-
Influence of cyclin type and dose on mitotic entry and progression in the early Drosophila embryo.J Cell Biol. 2009 Mar 9;184(5):639-46. doi: 10.1083/jcb.200810012. J Cell Biol. 2009. PMID: 19273612 Free PMC article.
-
Exploring the role of Drosophila cyclin A in the regulation of S phase.Cold Spring Harb Symp Quant Biol. 1991;56:465-75. doi: 10.1101/sqb.1991.056.01.053. Cold Spring Harb Symp Quant Biol. 1991. PMID: 1840259 No abstract available.
-
Synergistic action of Drosophila cyclins A and B during the G2-M transition.EMBO J. 1993 Jan;12(1):65-74. doi: 10.1002/j.1460-2075.1993.tb05632.x. EMBO J. 1993. PMID: 8428595 Free PMC article.
-
Cyclins and the G2/M transition.Cancer Surv. 1997;29:47-73. Cancer Surv. 1997. PMID: 9338096 Review.
-
All aboard the cyclin train: subcellular trafficking of cyclins and their CDK partners.Trends Cell Biol. 1999 Jun;9(6):207-10. doi: 10.1016/s0962-8924(99)01577-9. Trends Cell Biol. 1999. PMID: 10354564 Review.
Cited by
-
Waves of Cdk1 Activity in S Phase Synchronize the Cell Cycle in Drosophila Embryos.Dev Cell. 2016 Aug 22;38(4):399-412. doi: 10.1016/j.devcel.2016.07.023. Dev Cell. 2016. PMID: 27554859 Free PMC article.
-
Interphase-arrested Drosophila embryos activate zygotic gene expression and initiate mid-blastula transition events at a low nuclear-cytoplasmic ratio.PLoS Biol. 2020 Oct 22;18(10):e3000891. doi: 10.1371/journal.pbio.3000891. eCollection 2020 Oct. PLoS Biol. 2020. PMID: 33090988 Free PMC article.
-
Timing the Drosophila Mid-Blastula Transition: A Cell Cycle-Centered View.Trends Genet. 2016 Aug;32(8):496-507. doi: 10.1016/j.tig.2016.05.006. Epub 2016 Jun 20. Trends Genet. 2016. PMID: 27339317 Free PMC article. Review.
-
Regulation of DNA Replication in Early Embryonic Cleavages.Genes (Basel). 2017 Jan 19;8(1):42. doi: 10.3390/genes8010042. Genes (Basel). 2017. PMID: 28106858 Free PMC article. Review.
-
Cyclin B3 is a mitotic cyclin that promotes the metaphase-anaphase transition.Curr Biol. 2015 Mar 16;25(6):811-816. doi: 10.1016/j.cub.2015.01.053. Epub 2015 Mar 5. Curr Biol. 2015. PMID: 25754637 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
