

The essential polysome-associated RNA-binding protein RBP42 targets mRNAs involved in Trypanosoma brucei energy metabolism
- PMID: 22966087
- PMCID: PMC3479388
- DOI: 10.1261/rna.033829.112
The essential polysome-associated RNA-binding protein RBP42 targets mRNAs involved in Trypanosoma brucei energy metabolism
Abstract
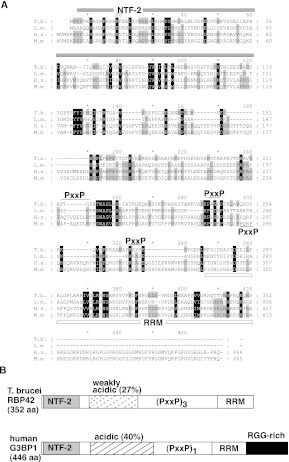
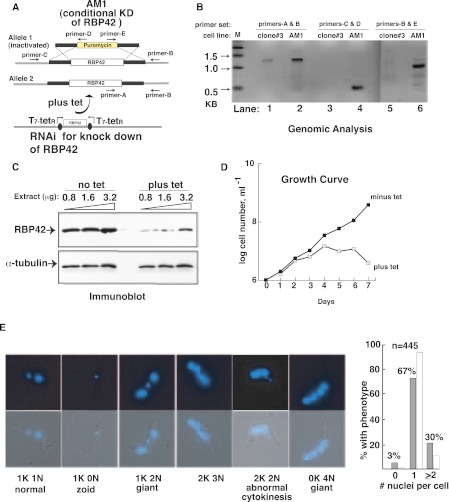
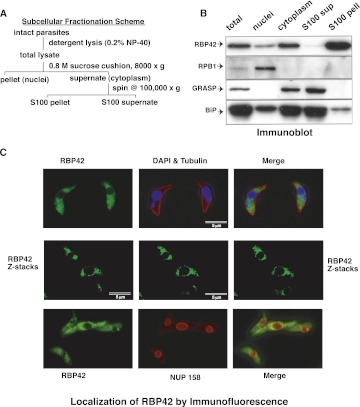
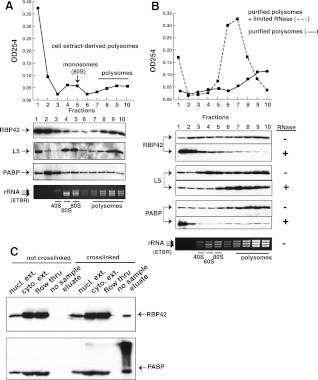
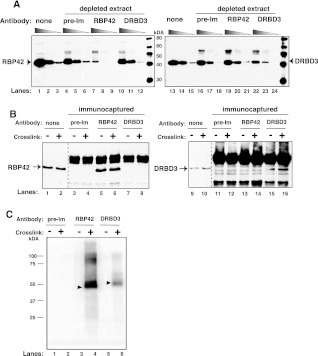
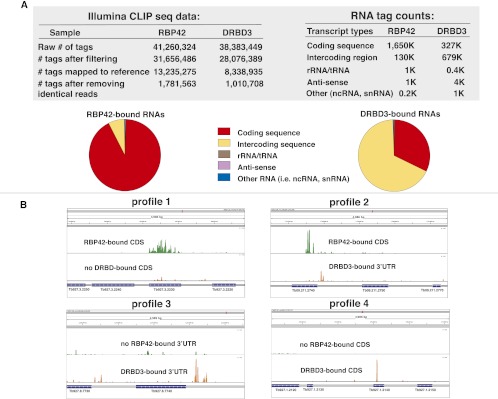
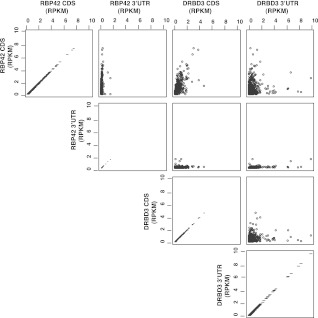
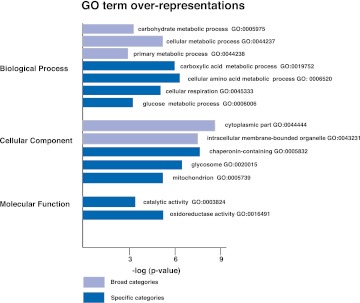
RNA-binding proteins that target mRNA coding regions are emerging as regulators of post-transcriptional processes in eukaryotes. Here we describe a newly identified RNA-binding protein, RBP42, which targets the coding region of mRNAs in the insect form of the African trypanosome, Trypanosoma brucei. RBP42 is an essential protein and associates with polysome-bound mRNAs in the cytoplasm. A global survey of RBP42-bound mRNAs was performed by applying HITS-CLIP technology, which captures protein-RNA interactions in vivo using UV light. Specific RBP42-mRNA interactions, as well as mRNA interactions with a known RNA-binding protein, were purified using specific antibodies. Target RNA sequences were identified and quantified using high-throughput RNA sequencing. Analysis revealed that RBP42 bound mainly within the coding region of mRNAs that encode proteins involved in cellular energy metabolism. Although the mechanism of RBP42's function is unclear at present, we speculate that RBP42 plays a critical role in modulating T. brucei energy metabolism.
Figures
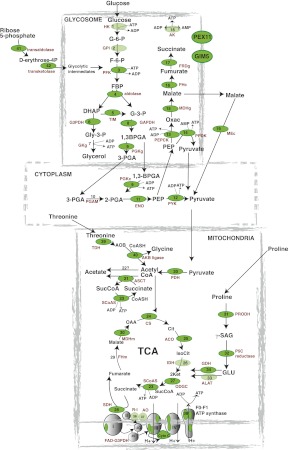
References
-
- Atlas R, Behar L, Elliott E, Ginzburg I 2004. The insulin-like growth factor mRNA binding-protein IMP-1 and the Ras-regulatory protein G3BP associate with tau mRNA and HuD protein in differentiated P19 neuronal cells. J Neurochem 89: 613–626 - PubMed
-
- Atlas R, Behar L, Sapoznik S, Ginzburg I 2007. Dynamic association with polysomes during P19 neuronal differentiation and an untranslated-region-dependent translation regulation of the tau mRNA by the tau mRNA-associated proteins IMP1, HuD, and G3BP1. J Neurosci Res 85: 173–183 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases