

Capsid protein expression and adeno-associated virus like particles assembly in Saccharomyces cerevisiae
- PMID: 22966759
- PMCID: PMC3539887
- DOI: 10.1186/1475-2859-11-124
Capsid protein expression and adeno-associated virus like particles assembly in Saccharomyces cerevisiae
Abstract
Background: The budding yeast Saccharomyces cerevisiae supports replication of many different RNA or DNA viruses (e.g. Tombusviruses or Papillomaviruses) and has provided means for up-scalable, cost- and time-effective production of various virus-like particles (e.g. Human Parvovirus B19 or Rotavirus). We have recently demonstrated that S. cerevisiae can form single stranded DNA AAV2 genomes starting from a circular plasmid. In this work, we have investigated the possibility to assemble AAV capsids in yeast.
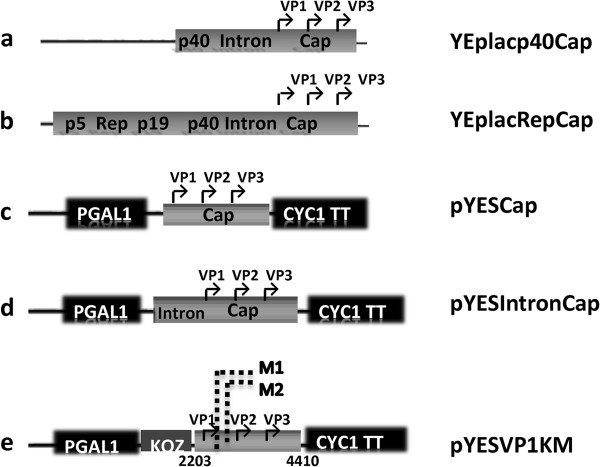
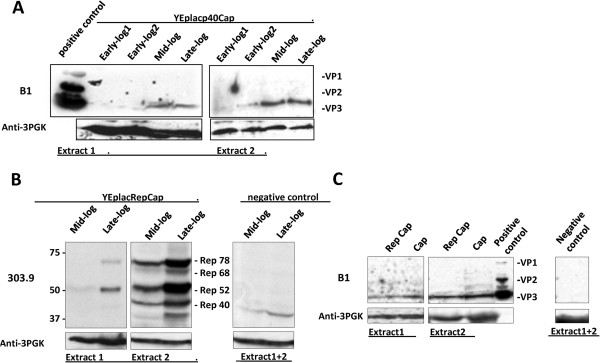
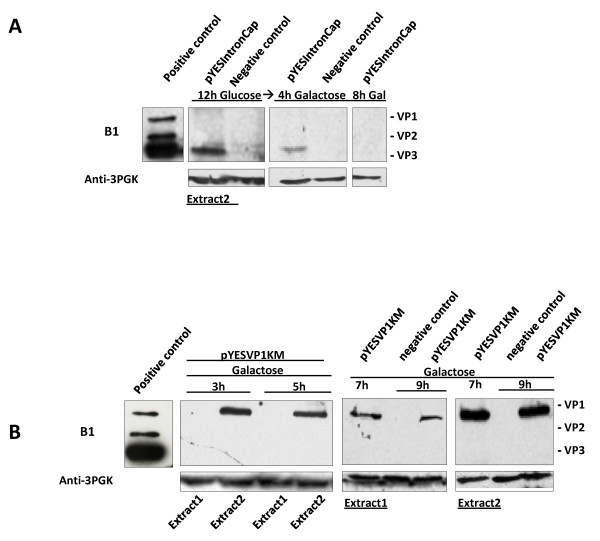
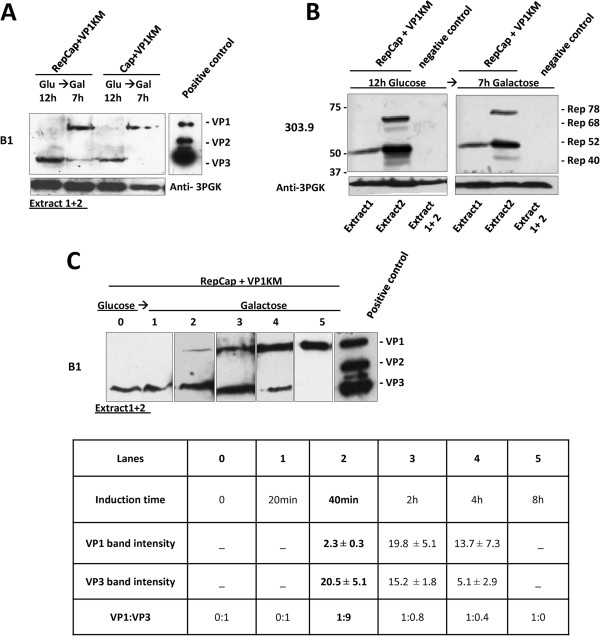
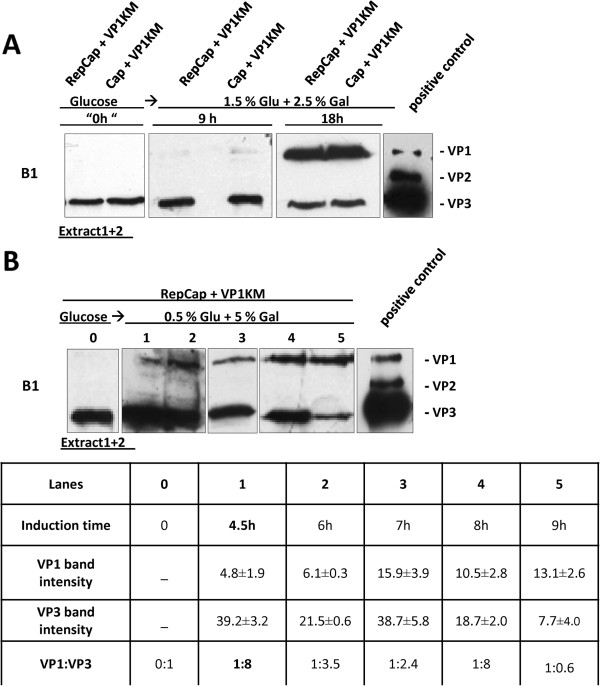
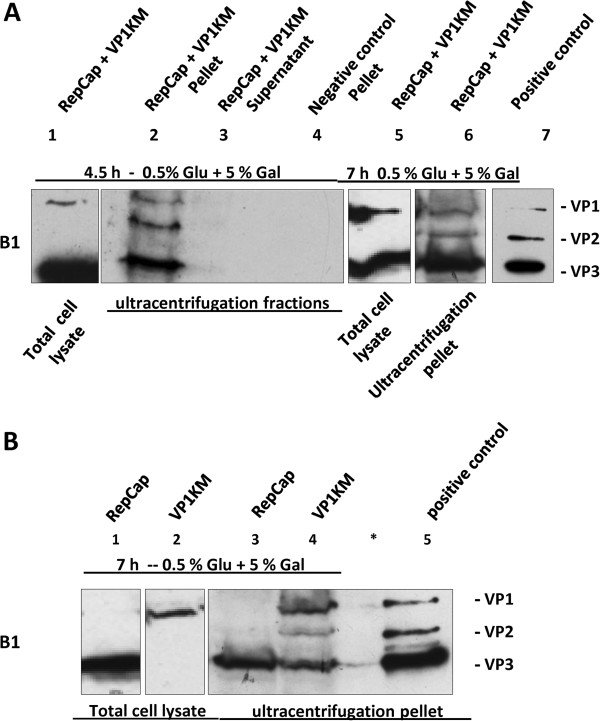
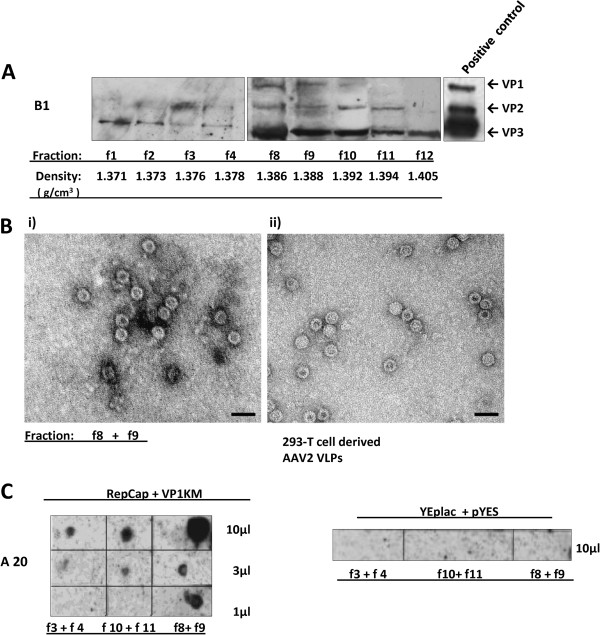
Results: To do this, at least two out of three AAV structural proteins, VP1 and VP3, have to be simultaneously expressed in yeast cells and their intracellular stoichiometry has to resemble the one found in the particles derived from mammalian or insect cells. This was achieved by stable co-transformation of yeast cells with two plasmids, one expressing VP3 from its natural p40 promoter and the other one primarily expressing VP1 from a modified AAV2 Cap gene under the control of the inducible yeast promoter Gal1. Among various induction strategies we tested, the best one to yield the appropriate VP1:VP3 ratio was 4.5 hour induction in the medium containing 0.5% glucose and 5% galactose. Following such induction, AAV virus like particles (VLPs) were isolated from yeast by two step ultracentrifugation procedure. The transmission electron microscopy analysis revealed that their morphology is similar to the empty capsids produced in human cells.
Conclusions: Taken together, the results show for the first time that yeast can be used to assemble AAV capsid and, therefore, as a genetic system to identify novel cellular factors involved in AAV biology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
