

Stat5 signaling specifies basal versus stress erythropoietic responses through distinct binary and graded dynamic modalities
- PMID: 22969412
- PMCID: PMC3433736
- DOI: 10.1371/journal.pbio.1001383
Stat5 signaling specifies basal versus stress erythropoietic responses through distinct binary and graded dynamic modalities
Abstract
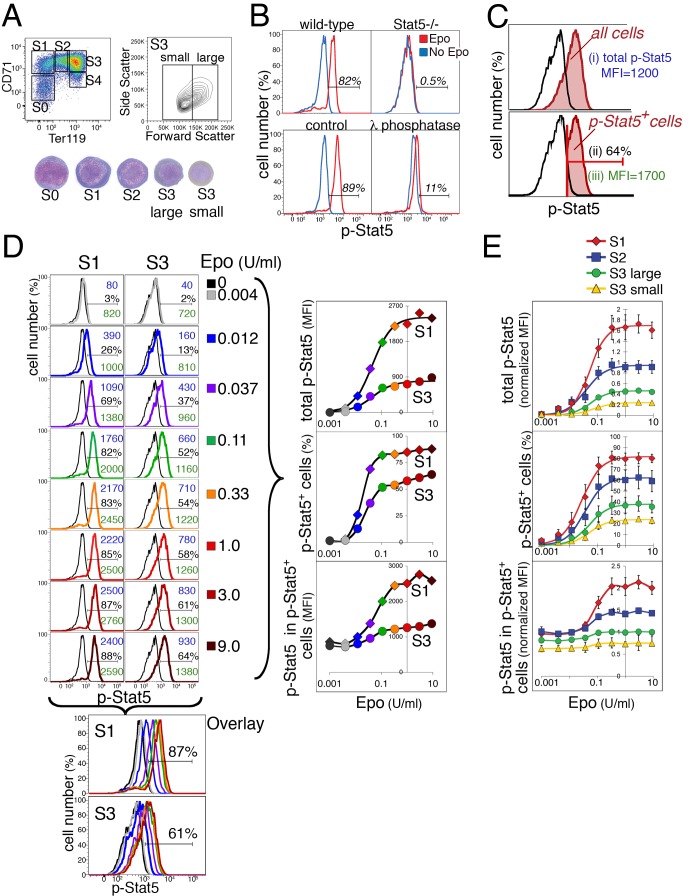
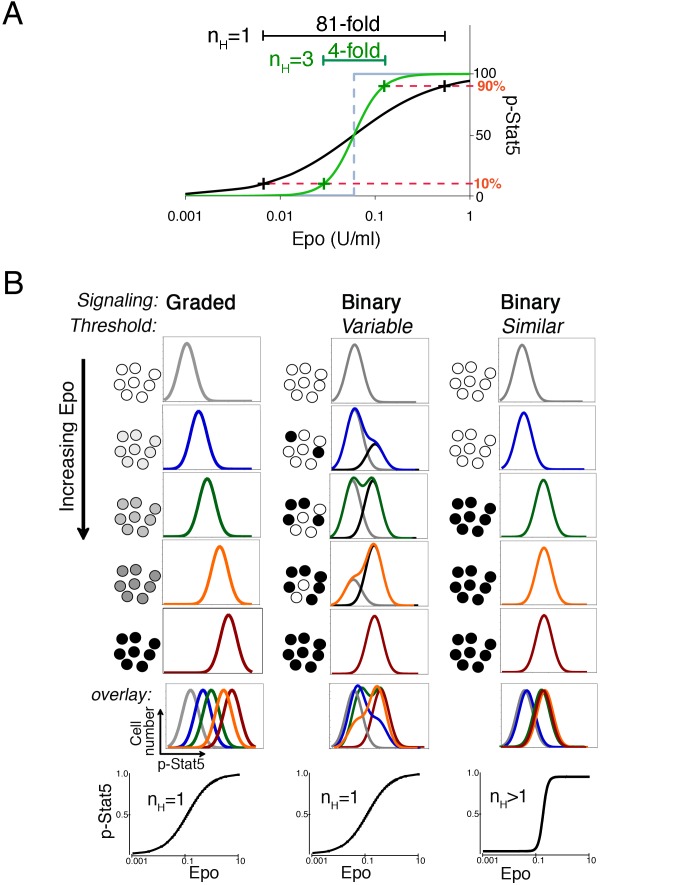
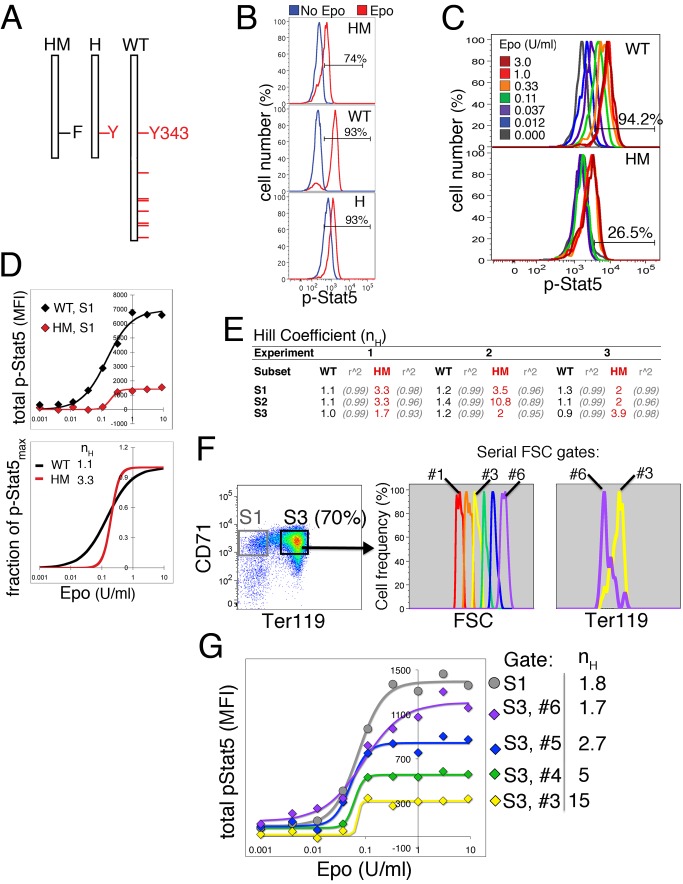
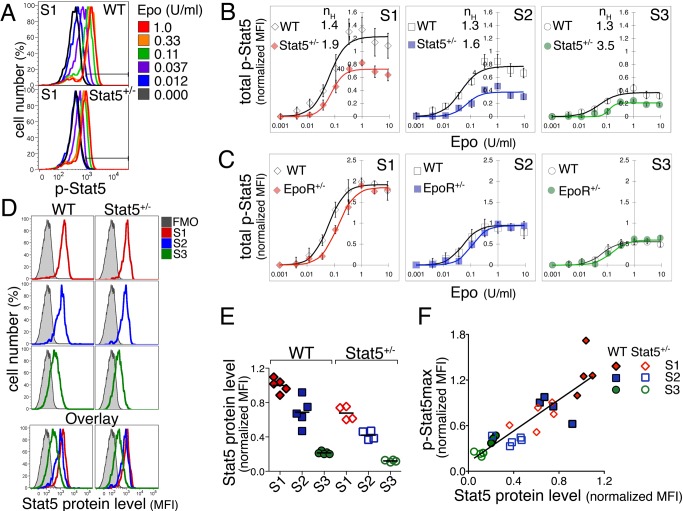
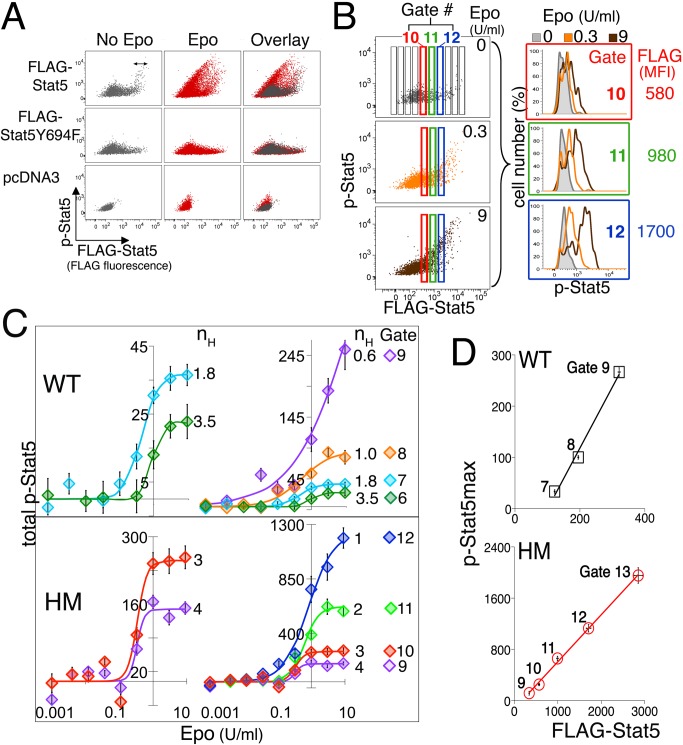
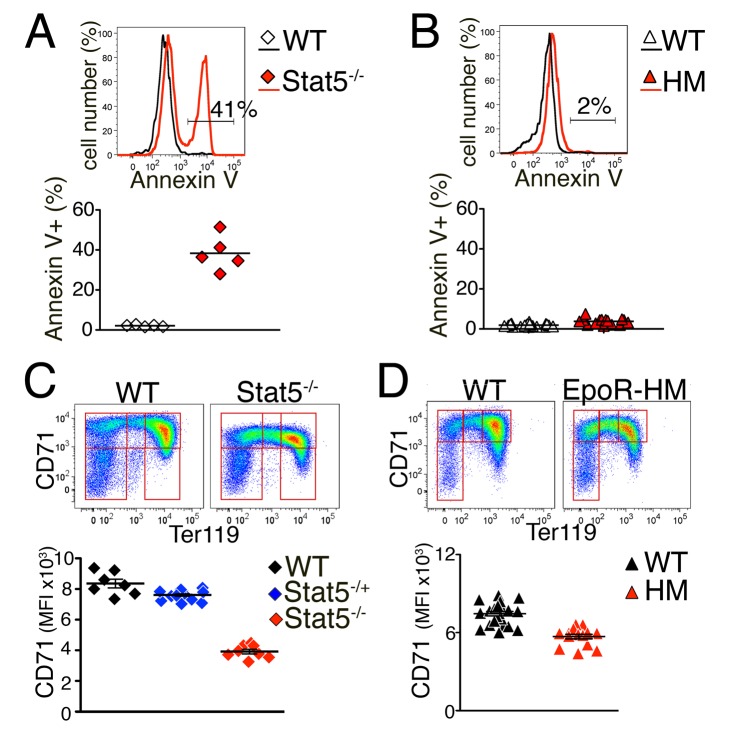
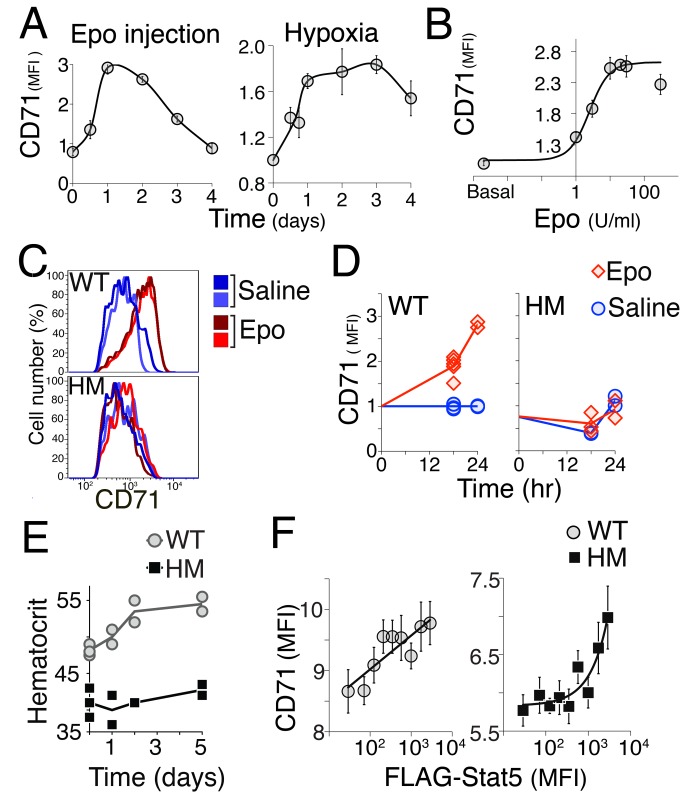
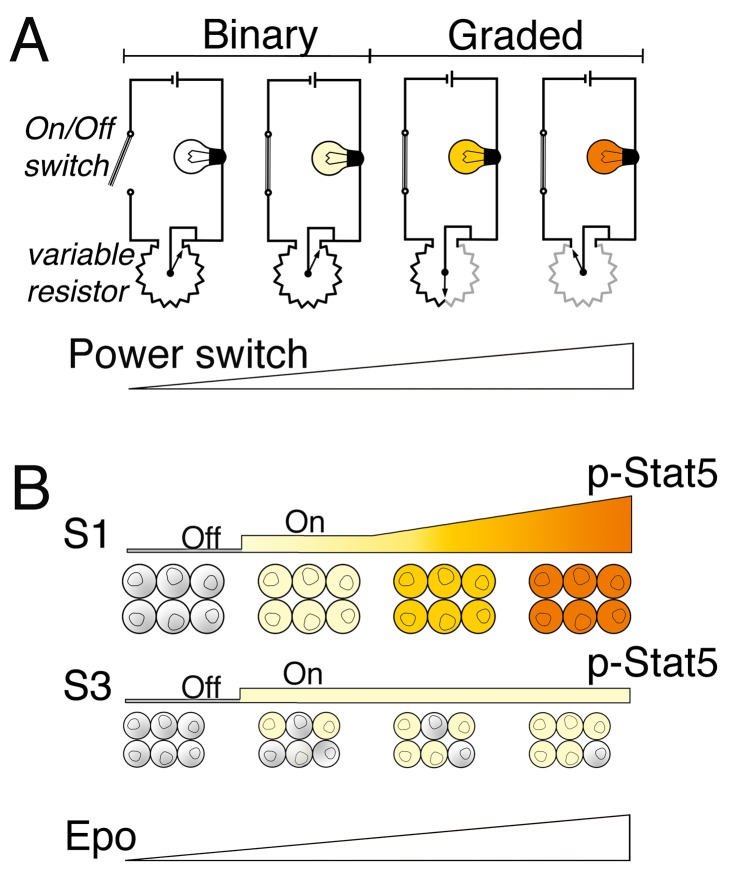
Erythropoietin (Epo)-induced Stat5 phosphorylation (p-Stat5) is essential for both basal erythropoiesis and for its acceleration during hypoxic stress. A key challenge lies in understanding how Stat5 signaling elicits distinct functions during basal and stress erythropoiesis. Here we asked whether these distinct functions might be specified by the dynamic behavior of the Stat5 signal. We used flow cytometry to analyze Stat5 phosphorylation dynamics in primary erythropoietic tissue in vivo and in vitro, identifying two signaling modalities. In later (basophilic) erythroblasts, Epo stimulation triggers a low intensity but decisive, binary (digital) p-Stat5 signal. In early erythroblasts the binary signal is superseded by a high-intensity graded (analog) p-Stat5 response. We elucidated the biological functions of binary and graded Stat5 signaling using the EpoR-HM mice, which express a "knocked-in" EpoR mutant lacking cytoplasmic phosphotyrosines. Strikingly, EpoR-HM mice are restricted to the binary signaling mode, which rescues these mice from fatal perinatal anemia by promoting binary survival decisions in erythroblasts. However, the absence of the graded p-Stat5 response in the EpoR-HM mice prevents them from accelerating red cell production in response to stress, including a failure to upregulate the transferrin receptor, which we show is a novel stress target. We found that Stat5 protein levels decline with erythroblast differentiation, governing the transition from high-intensity graded signaling in early erythroblasts to low-intensity binary signaling in later erythroblasts. Thus, using exogenous Stat5, we converted later erythroblasts into high-intensity graded signal transducers capable of eliciting a downstream stress response. Unlike the Stat5 protein, EpoR expression in erythroblasts does not limit the Stat5 signaling response, a non-Michaelian paradigm with therapeutic implications in myeloproliferative disease. Our findings show how the binary and graded modalities combine to generate high-fidelity Stat5 signaling over the entire basal and stress Epo range. They suggest that dynamic behavior may encode information during STAT signal transduction.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
How a blood cell intracellular signal keeps blood flowing.PLoS Biol. 2012 Aug;10(8):e1001385. doi: 10.1371/journal.pbio.1001385. Epub 2012 Aug 28. PLoS Biol. 2012. PMID: 22952432 Free PMC article. No abstract available.
References
-
- D'Andrea AD, Lodish HF, Wong GG (1989) Expression cloning of the murine erythropoietin receptor. Cell 57: 277–285. - PubMed
-
- Wu H, Liu X, Jaenisch R, Lodish HF (1995) Generation of committed erythroid BFU-E and CFU-E progenitors does not require erythropoietin or the erythropoietin receptor. Cell 83: 59–67. - PubMed
-
- Koulnis M, Liu Y, Hallstrom K, Socolovsky M (2011) Negative autoregulation by Fas stabilizes adult erythropoiesis and accelerates its stress response. PLoS One 6: e21192 doi:10.1371/journal.pone.0021192. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
