

A chick model of retinal detachment: cone rich and novel
- PMID: 22970190
- PMCID: PMC3435403
- DOI: 10.1371/journal.pone.0044257
A chick model of retinal detachment: cone rich and novel
Abstract
Background: Development of retinal detachment models in small animals can be difficult and expensive. Here we create and characterize a novel, cone-rich retinal detachment (RD) model in the chick.
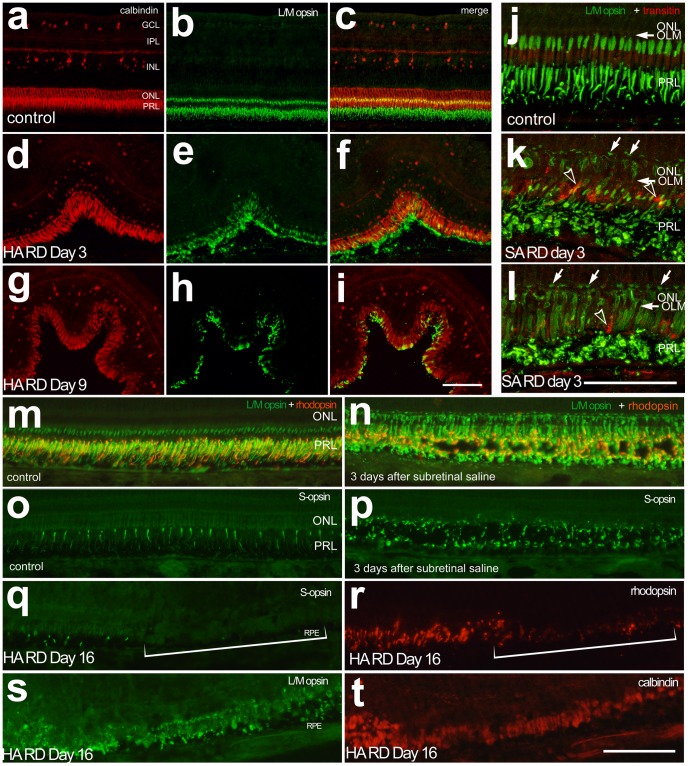
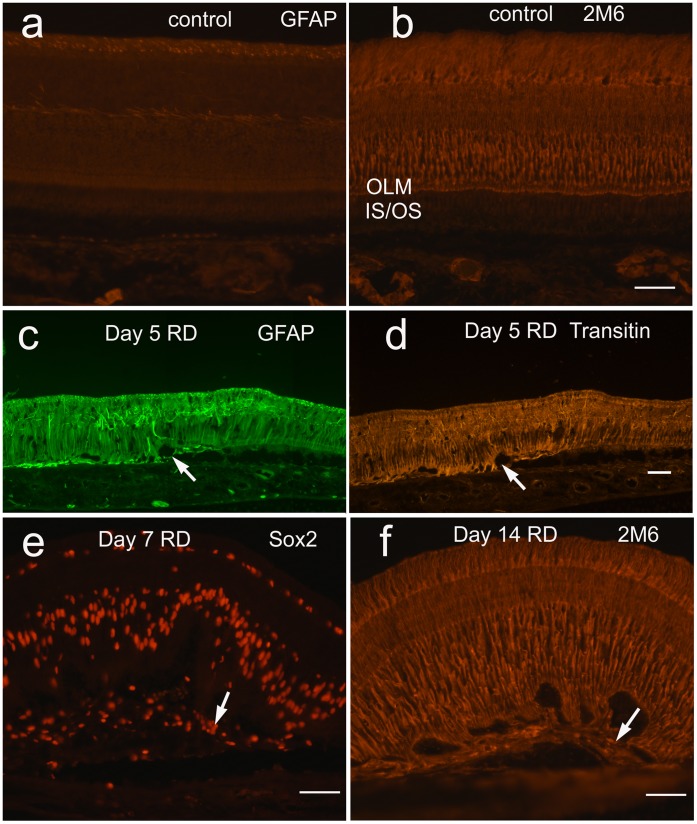
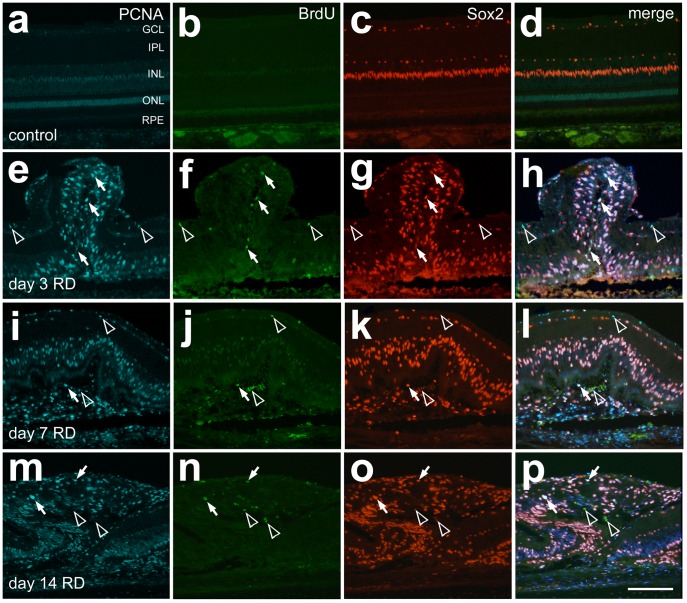
Methodology/principal findings: Retinal detachments were created in chicks between postnatal days 7 and 21 by subretinal injections of either saline (SA) or hyaluronic acid (HA). Injections were performed through a dilated pupil with observation via surgical microscope, using the fellow eye as a control. Immunohistochemical analyses were performed at days 1, 3, 7, 10 and 14 after retinal detachment to evaluate the cellular responses of photoreceptors, Müller glia, microglia and nonastrocytic inner retinal glia (NIRG). Cell proliferation was detected with bromodeoxyuridine (BrdU)-incorporation and by the expression of proliferating cell nuclear antigen (PCNA). Cell death was detected with terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL). As in mammalian models of RD, there is shortening of photoreceptor outer segments and mis-trafficking of photoreceptor opsins in areas of RD. Photoreceptor cell death was maximal 1 day after RD, but continued until 14 days after RD. Müller glia up-regulated glial fibriliary acidic protein (GFAP), proliferated, showed interkinetic nuclear migration, and migrated to the subretinal space in areas of detachment. Microglia became reactive; they up-regulated CD45, acquired amoeboid morphology, and migrated toward outer retina in areas of RD. Reactive NIRG cells accumulated in detached areas.
Conclusions/significance: Subretinal injections of SA or HA in the chick eye successfully produced retinal detachments and cellular responses similar to those seen in standard mammalian models. Given the relatively large eye size, and considering the low cost, the chick model of RD offers advantages for high-throughput studies.
Conflict of interest statement
Figures




References
-
- Agrawal RN, He S, Spee C, Cui JZ, Ryan SJ, et al. (2007) In vivo models of proliferative vitreoretinopathy. Nat Protoc 2: 67–77. - PubMed
-
- Mandava N, Blackburn P, Paul DB, Wilson MW, Read SB, et al. (2002) Ribozyme to proliferating cell nuclear antigen to treat proliferative vitreoretinopathy. Invest Ophthalmol Vis Sci 43: 3338–3348. - PubMed
-
- Schiff WM, Hwang JC, Ober MD, Olson JL, Dhrami-Gavazi E, et al. (2007) Safety and efficacy assessment of chimeric ribozyme to proliferating cell nuclear antigen to prevent recurrence of proliferative vitreoretinopathy. Arch Ophthalmol 125: 1161–1167. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
