

Cyclic AMP dynamics in the pancreatic β-cell
- PMID: 22970724
- PMCID: PMC3497220
- DOI: 10.3109/03009734.2012.724732
Cyclic AMP dynamics in the pancreatic β-cell
Abstract
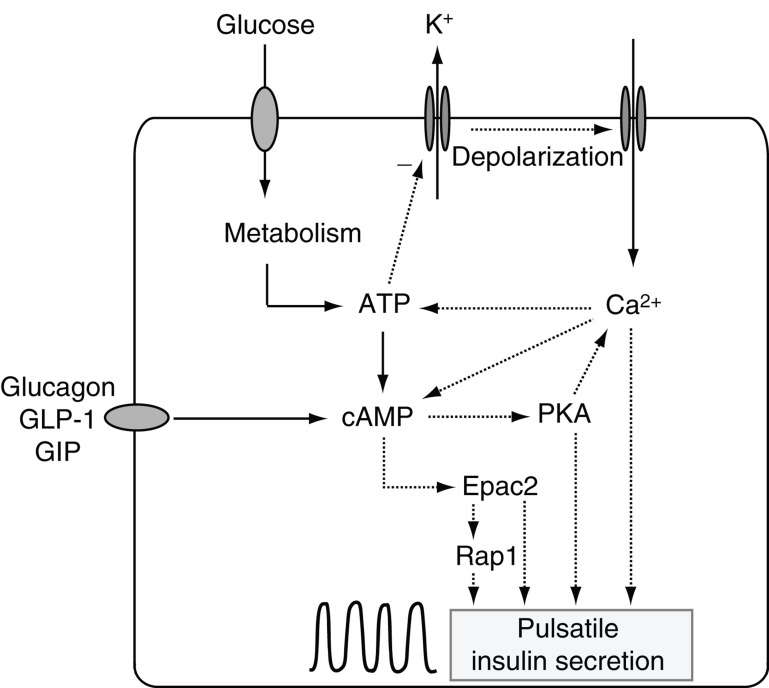
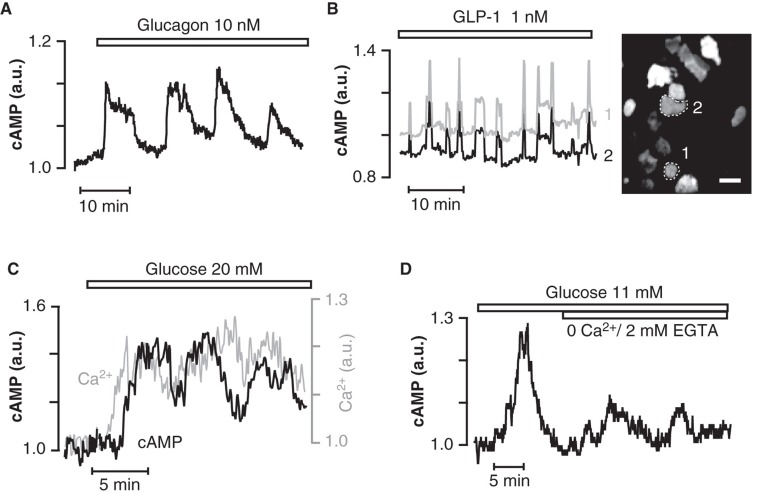
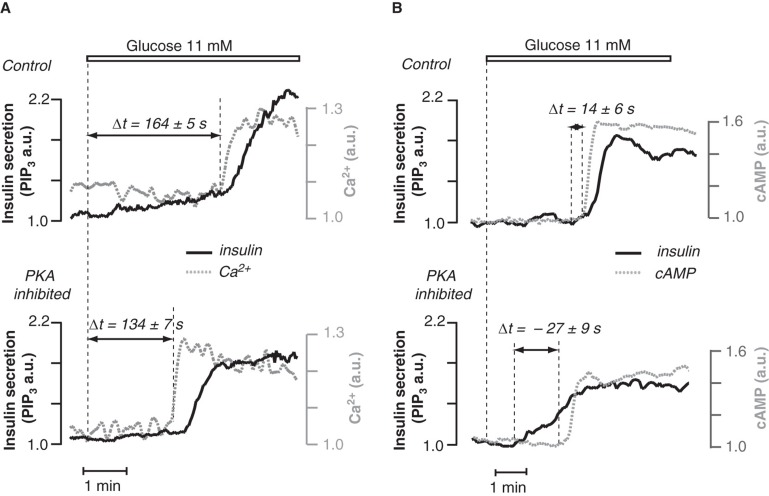
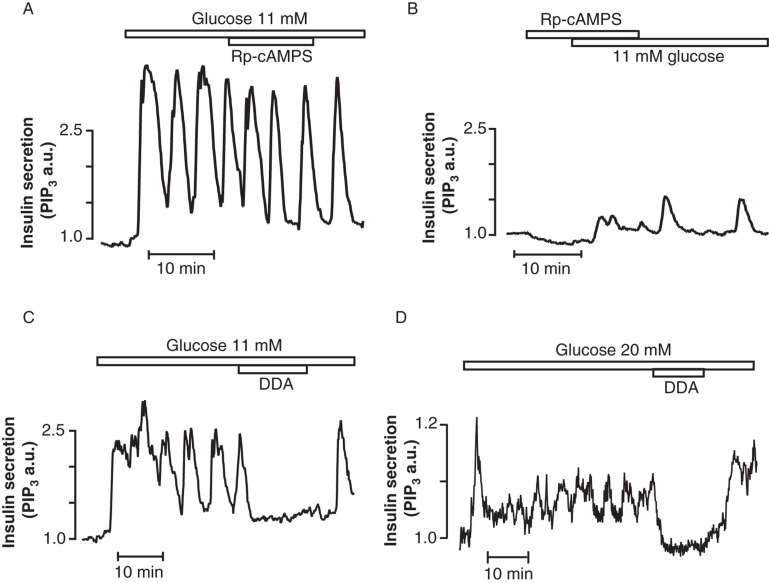
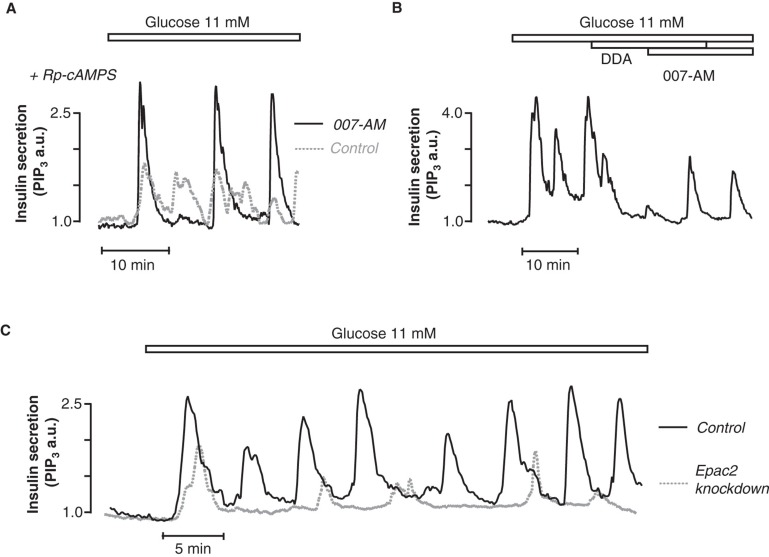
Insulin secretion from pancreatic β-cells is tightly regulated by glucose and other nutrients, hormones, and neural factors. The exocytosis of insulin granules is triggered by an elevation of the cytoplasmic Ca(2+) concentration ([Ca(2+)](i)) and is further amplified by cyclic AMP (cAMP). Cyclic AMP is formed primarily in response to glucoincretin hormones and other G(s)-coupled receptor agonists, but generation of the nucleotide is critical also for an optimal insulin secretory response to glucose. Nutrient and receptor stimuli trigger oscillations of the cAMP concentration in β-cells. The oscillations arise from variations in adenylyl cyclase-mediated cAMP production and phosphodiesterase-mediated degradation, processes controlled by factors like cell metabolism and [Ca(2+)](i). Protein kinase A and the guanine nucleotide exchange factor Epac2 mediate the actions of cAMP in β-cells and operate at multiple levels to promote exocytosis and pulsatile insulin secretion. The cAMP signaling system contains important targets for pharmacological improvement of insulin secretion in type 2 diabetes.
Figures
References
-
- Pørksen N. The in vivo regulation of pulsatile insulin secretion. Diabetologia. 2002;45:3–20. - PubMed
-
- Schmitz O, Rungby J, Edge L, Juhl CB. On high-frequency insulin oscillations. Ageing Res Rev. 2008;7:301–5. - PubMed
-
- Tengholm A, Gylfe E. Oscillatory control of insulin secretion. Mol Cell Endocrinol. 2009;297:58–72. - PubMed
-
- Gilon P, Ravier MA, Jonas JC, Henquin JC. Control mechanisms of the oscillations of insulin secretion in vitro and in vivo. Diabetes. 2002;51:S144–51. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous