

Activity-dependent regulation of retinogeniculate signaling by metabotropic glutamate receptors
- PMID: 22973005
- PMCID: PMC3462222
- DOI: 10.1523/JNEUROSCI.0687-12.2012
Activity-dependent regulation of retinogeniculate signaling by metabotropic glutamate receptors
Abstract
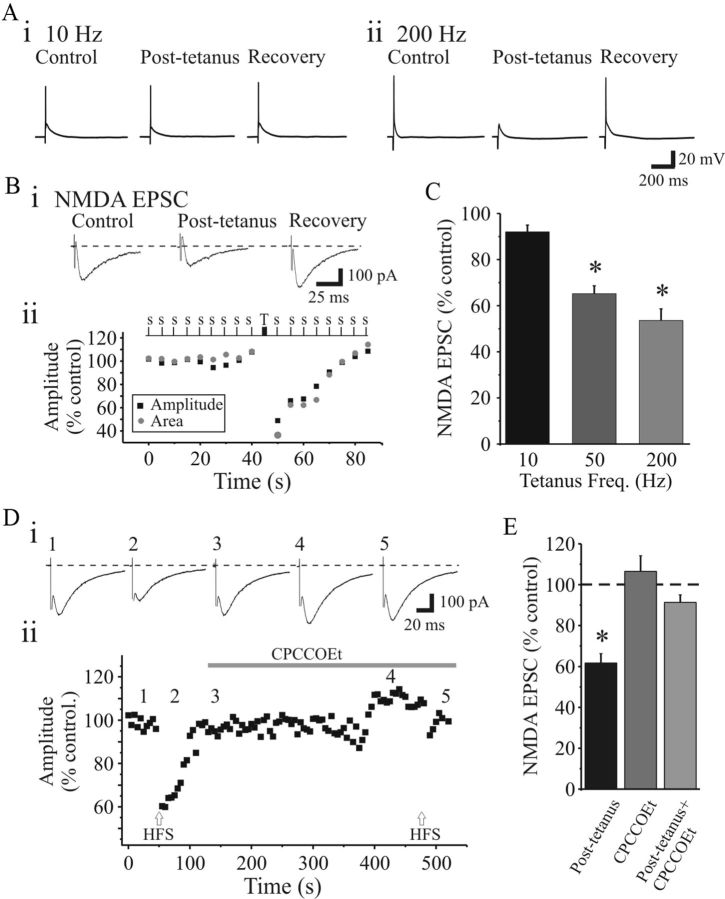
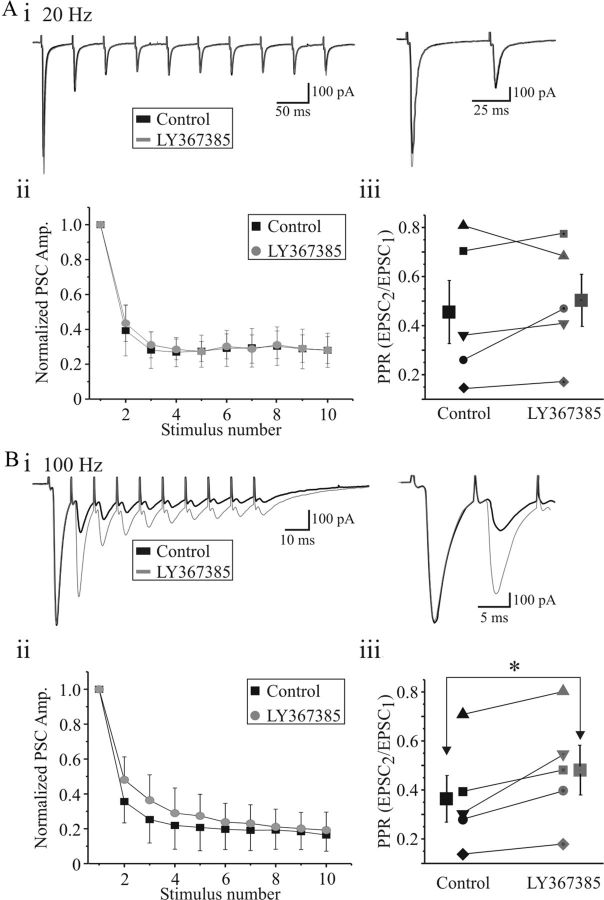
Thalamocortical neurons in dorsal lateral geniculate nucleus (dLGN) dynamically convey visual information from retina to the neocortex. Activation of metabotropic glutamate receptors (mGluRs) exerts multiple effects on neural integration in dLGN; however, their direct influence on the primary sensory input, namely retinogeniculate afferents, is unknown. In the present study, we found that pharmacological or synaptic activation of type 1 mGluRs (mGluR(1)s) significantly depresses glutamatergic retinogeniculate excitation in rat thalamocortical neurons. Pharmacological activation of mGluR(1)s attenuates excitatory synaptic responses in thalamocortical neurons at a magnitude sufficient to decrease suprathreshold output of these neurons. The reduction in both NMDA and AMPA receptor-dependent synaptic responses results from a presynaptic reduction in glutamate release from retinogeniculate terminals. The suppression of retinogeniculate synaptic transmission and dampening of thalamocortical output was mimicked by tetanic activation of retinogeniculate afferent in a frequency-dependent manner that activated mGluR(1)s. Retinogeniculate excitatory synaptic transmission was also suppressed by the glutamate transport blocker TBOA (dl-threo-β-benzyloxyaspartic acid), suggesting that mGluR(1)s were activated by glutamate spillover. The data indicate that presynaptic mGluR(1) contributes to an activity-dependent mechanism that regulates retinogeniculate excitation and therefore plays a significant role in the thalamic gating of visual information.
Figures
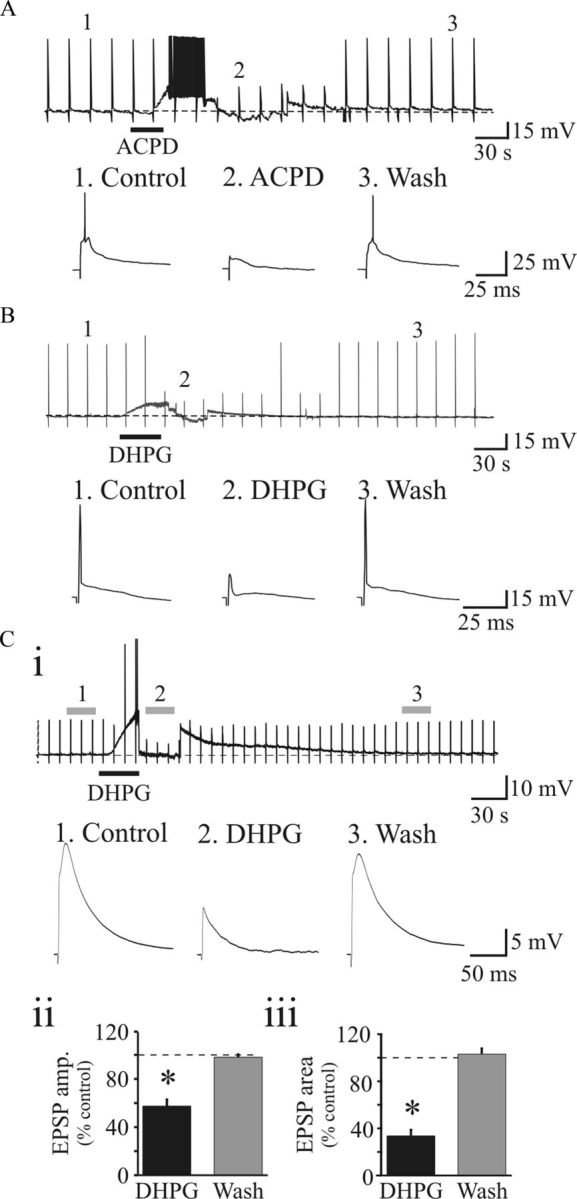
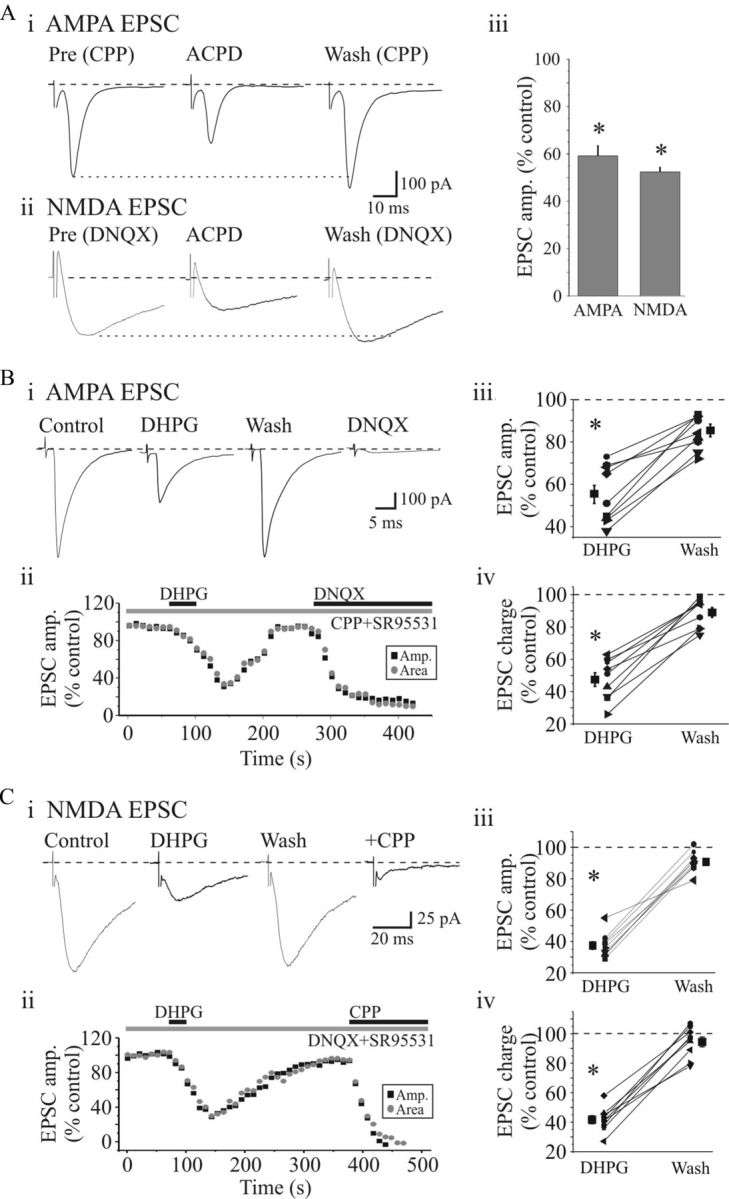
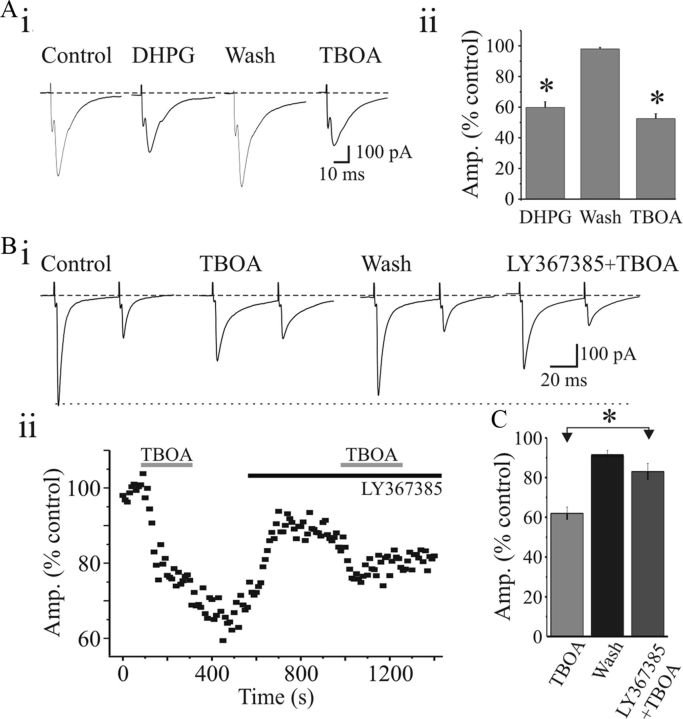
 ). Population data reveal a significant suppression of AMPAR EPSC amplitude (Biii) and charge (Biv) by DHPG that recovers near baseline level following washout. Ci, Representative NMDAR-dependent EPSCs recorded in presence of DNQX and SR95531. DHPG (25 μ) that recovers to baseline levels following washout. NMDAR EPSCs were attenuated by
). Population data reveal a significant suppression of AMPAR EPSC amplitude (Biii) and charge (Biv) by DHPG that recovers near baseline level following washout. Ci, Representative NMDAR-dependent EPSCs recorded in presence of DNQX and SR95531. DHPG (25 μ) that recovers to baseline levels following washout. NMDAR EPSCs were attenuated by 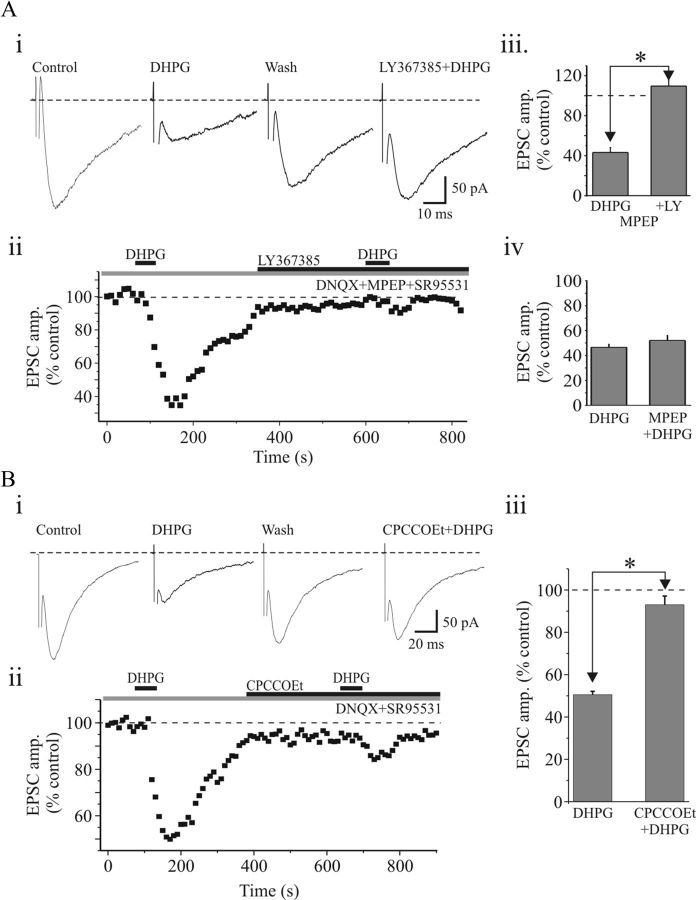
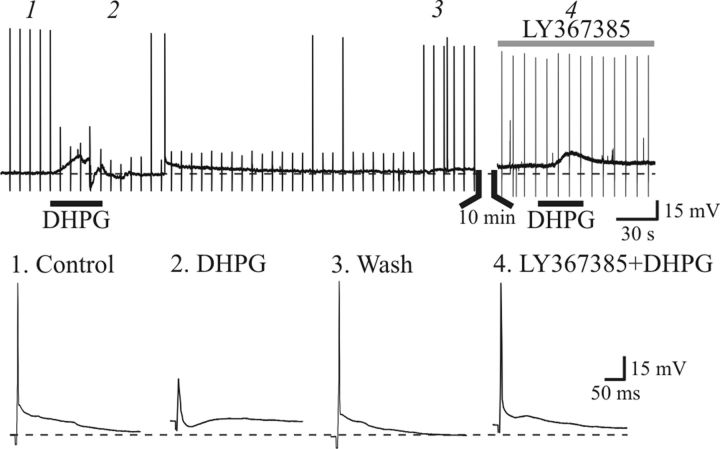
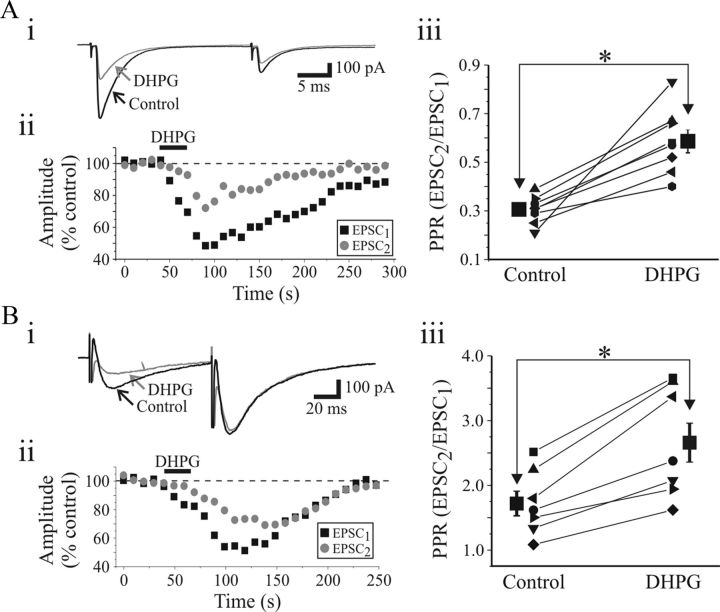 ). Aiii, Population data reveal that paired-pulse ratio (PPR = EPSC2/EPSC1) is significantly increased after DHPG application. Bi, In a different neuron, paired-pulse stimulation of OT (75 ms ISI) produces a facilitation of NMDAR EPSCs. DHPG (25 μ). Biii, Population data reveal that DHPG significantly increases the PPR. * p < 0.01.
). Aiii, Population data reveal that paired-pulse ratio (PPR = EPSC2/EPSC1) is significantly increased after DHPG application. Bi, In a different neuron, paired-pulse stimulation of OT (75 ms ISI) produces a facilitation of NMDAR EPSCs. DHPG (25 μ). Biii, Population data reveal that DHPG significantly increases the PPR. * p < 0.01.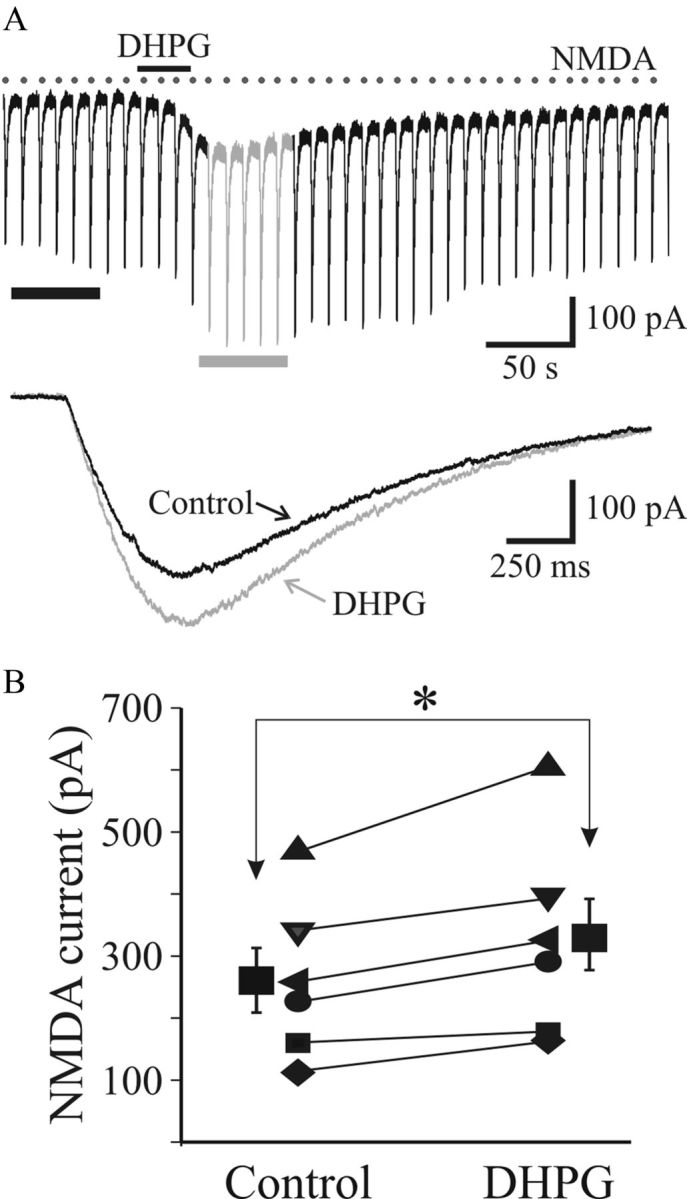
References
-
- Alexander GM, Godwin DW. Presynaptic inhibition of corticothalamic feedback by metabotropic glutamate receptors. J Neurophysiol. 2005;94:163–175. - PubMed
-
- Arnth-Jensen N, Jabaudon D, Scanziani M. Cooperation between independent hippocampal synapses is controlled by glutamate uptake. Nat Neurosci. 2002;5:325–331. - PubMed
-
- Chen C, Regehr WG. Developmental remodeling of the retinogeniculate synapse. Neuron. 2000;28:955–966. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources