

The different ligand-binding modes of relaxin family peptide receptors RXFP1 and RXFP2
- PMID: 22973049
- PMCID: PMC3487622
- DOI: 10.1210/me.2012-1188
The different ligand-binding modes of relaxin family peptide receptors RXFP1 and RXFP2
Abstract
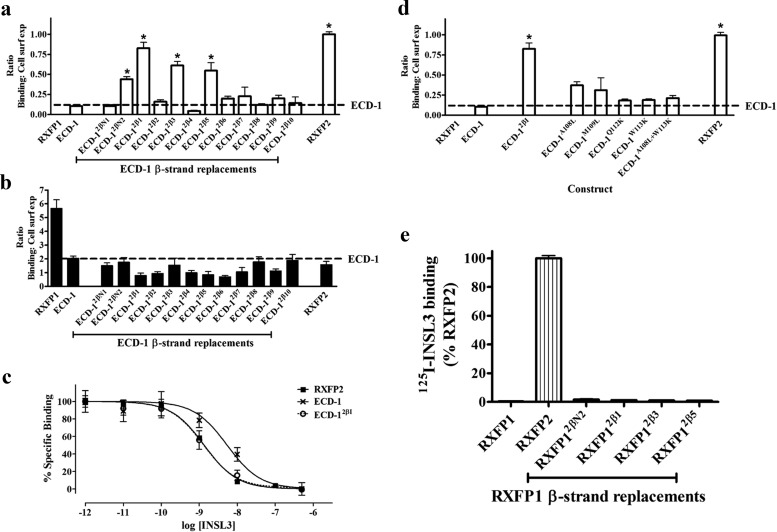
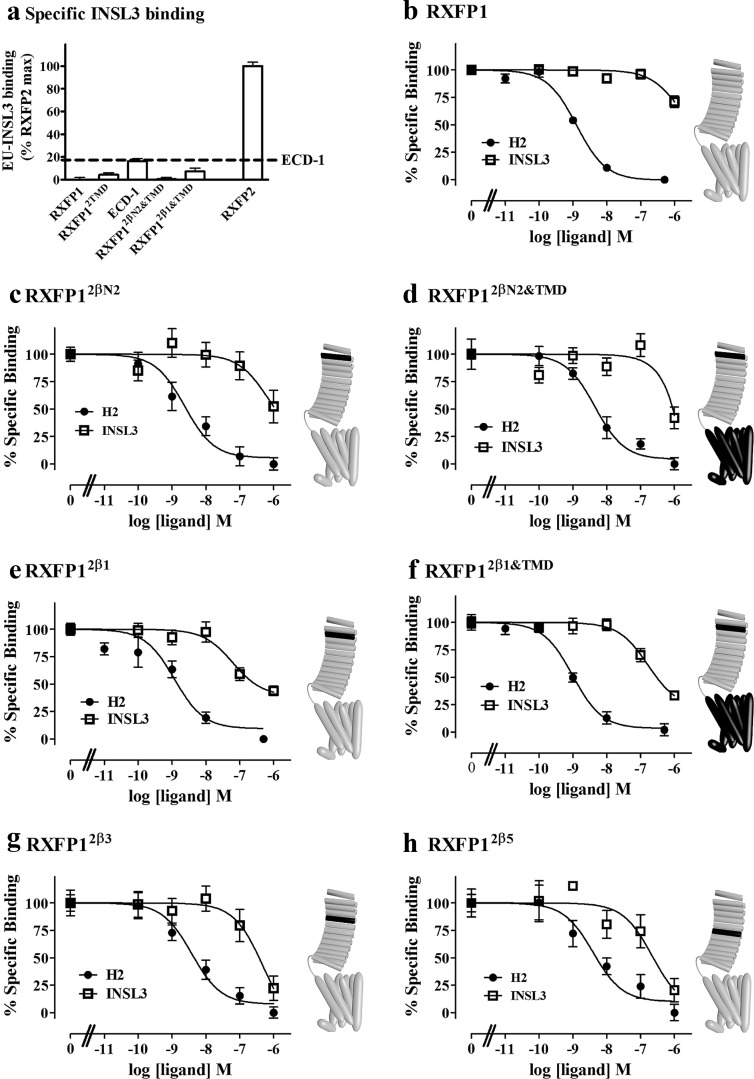
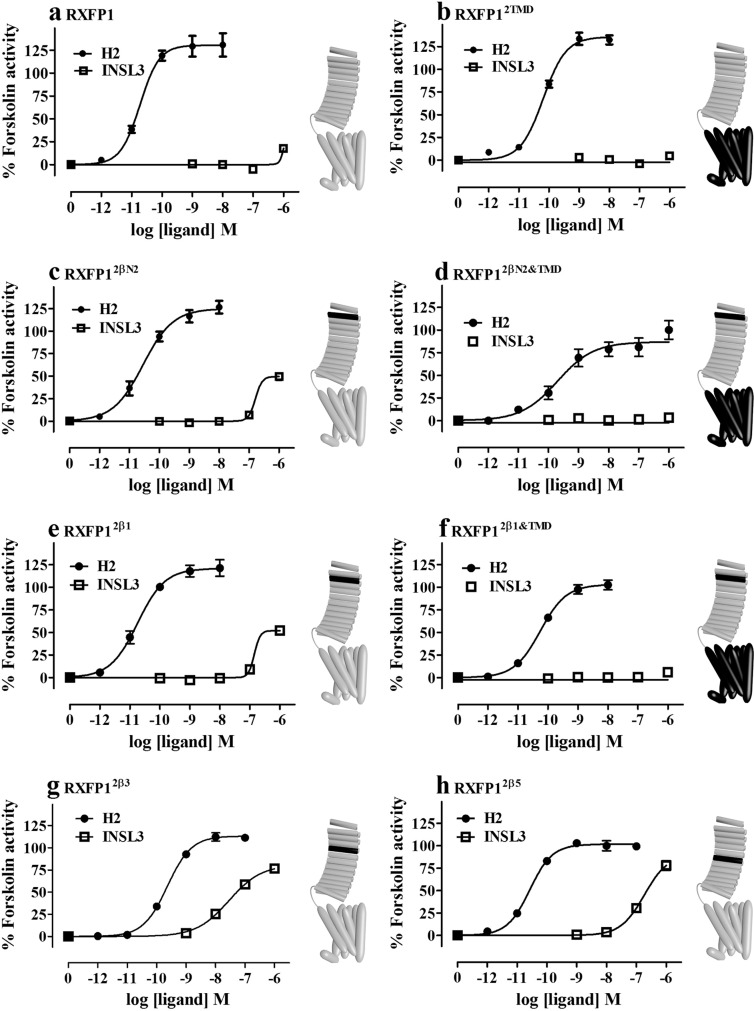
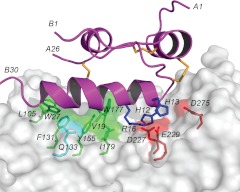
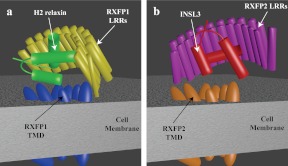
Relaxin and insulin-like peptide 3 (INSL3) are peptide hormones with a number of important physiological roles in reproduction, regulation of extracellular matrix turnover, and cardiovascular function. Relaxin and INSL3 mediate their actions through the closely related G-protein coupled receptors, relaxin family peptide receptors 1 and 2 (RXFP1 and RXFP2), respectively. These receptors have large extracellular domains (ECD) that contain high-affinity ligand-binding sites within their 10 leucine-rich repeat (LRR)-containing modules. Although relaxin can bind and activate both RXFP1 and RXFP2, INSL3 can only bind and activate RXFP2. To investigate whether this difference is related to the nature of the high-affinity ECD binding site or to differences in secondary binding sites involving the receptor transmembrane (TM) domain, we created a suite of constructs with RXFP1/2 chimeric ECD attached to single TM helices. We show that by changing as little as one LRR, representing four amino acid substitutions, we were able to engineer a high-affinity INSL3-binding site into the ECD of RXFP1. Molecular modeling of the INSL3-RXFP2 interaction based on extensive experimental data highlights the differences in the binding mechanisms of relaxin and INSL3 to the ECD of their cognate receptors. Interestingly, when the engineered RXFP1/2 ECD were introduced into full-length RXFP1 constructs, INSL3 exhibited only low affinity and efficacy on these receptors. These results highlight critical differences both in the ECD binding and in the coordination of the ECD-binding site with the TM domain, and provide new mechanistic insights into the binding and activation events of RXFP1 and RXFP2 by their native hormone ligands.
Figures

References
-
- Bathgate RAD, Hsueh AJ, Sherwood OD. 2006. Physiology and molecular biology of the relaxin peptide family. In: (Secondary Bathgate RAD, Hsueh AJ, Sherwood OD.) (Neill JD, ed. Physiology of reproduction. 3rd ed San Diego: Elsevier; 679–770
-
- Bathgate RA, Ivell R, Sanborn BM, Sherwood OD, Summers RJ. 2006. International Union of Pharmacology: recommendations for the nomenclature of receptors for relaxin family peptides. Pharmacol Rev 58:7–31 - PubMed
-
- Hsu SY, Liang SG, Hsueh AJ. 1998. Characterization of two LGR genes homologous to gonadotropin and thyrotropin receptors with extracellular leucine-rich repeats and a G protein-coupled, seven-transmembrane region. Mol Endocrinol 12:1830–1845 - PubMed
-
- Scott DJ, Layfield S, Yan Y, Sudo S, Hsueh AJ, Tregear GW, Bathgate RA. 2006. Characterization of novel splice variants of LGR7 and LGR8 reveals that receptor signaling is mediated by their unique LDLa modules. J Biol Chem 281:34942–34954 - PubMed
-
- Hopkins EJ, Layfield S, Ferraro T, Bathgate RA, Gooley PR. 2007. The NMR solution structure of the relaxin (RXFP1) receptor lipoprotein receptor class A module and identification of key residues in the N-terminal region of the module that mediate receptor activation. J Biol Chem 282:4172–4184 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
