

Identification of a dynamic mitochondrial protein complex driving cholesterol import, trafficking, and metabolism to steroid hormones
- PMID: 22973050
- PMCID: PMC5416962
- DOI: 10.1210/me.2012-1159
Identification of a dynamic mitochondrial protein complex driving cholesterol import, trafficking, and metabolism to steroid hormones
Abstract
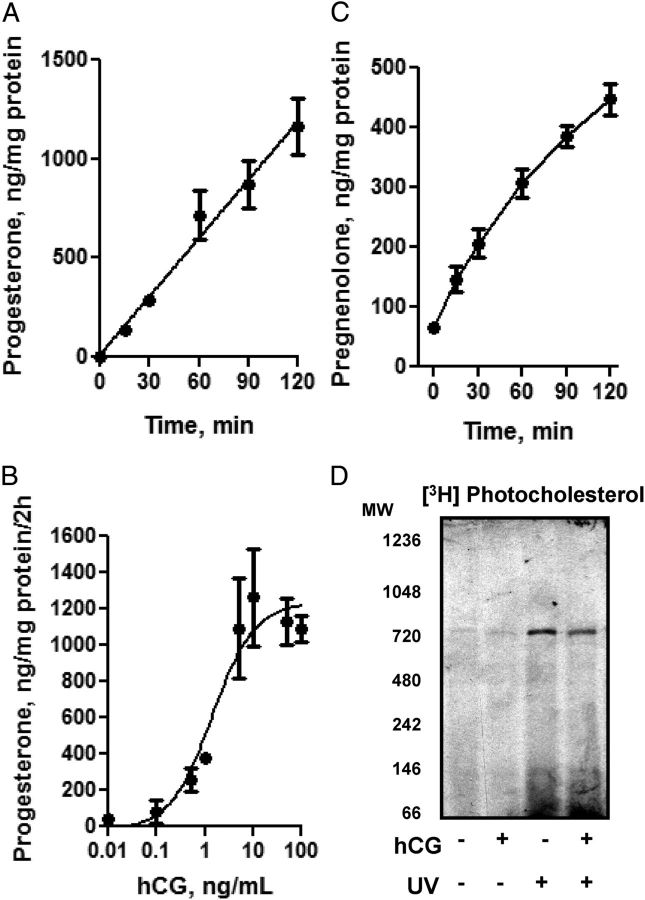
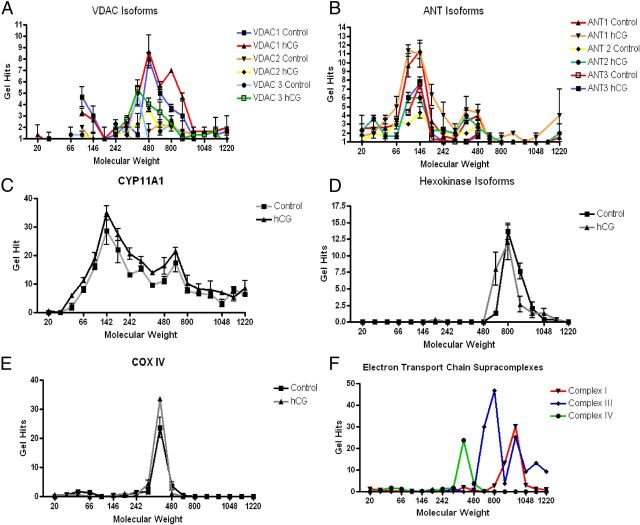
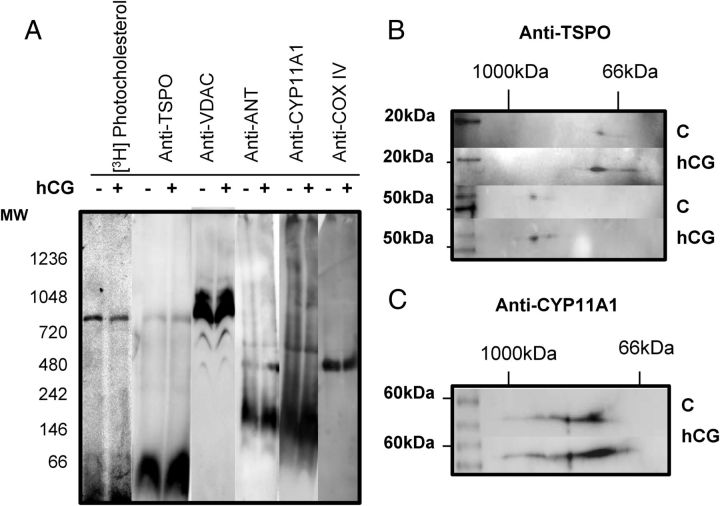
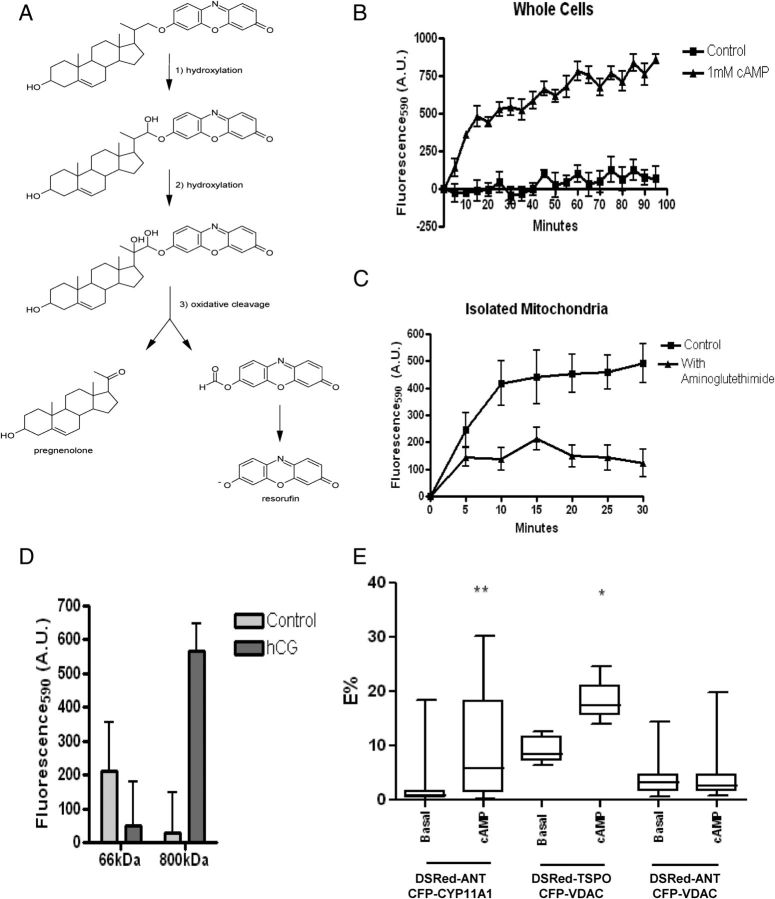
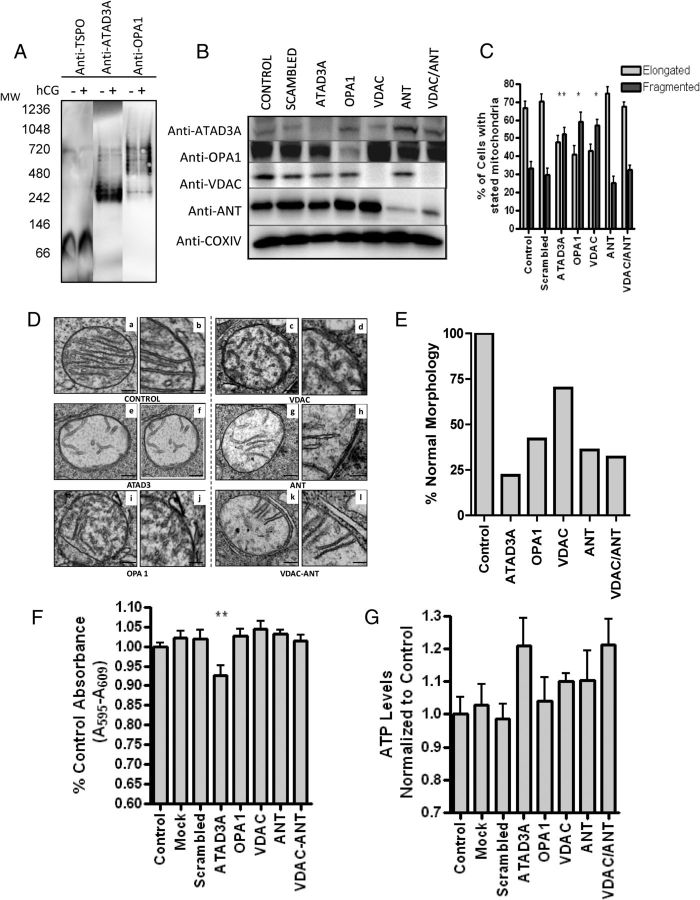
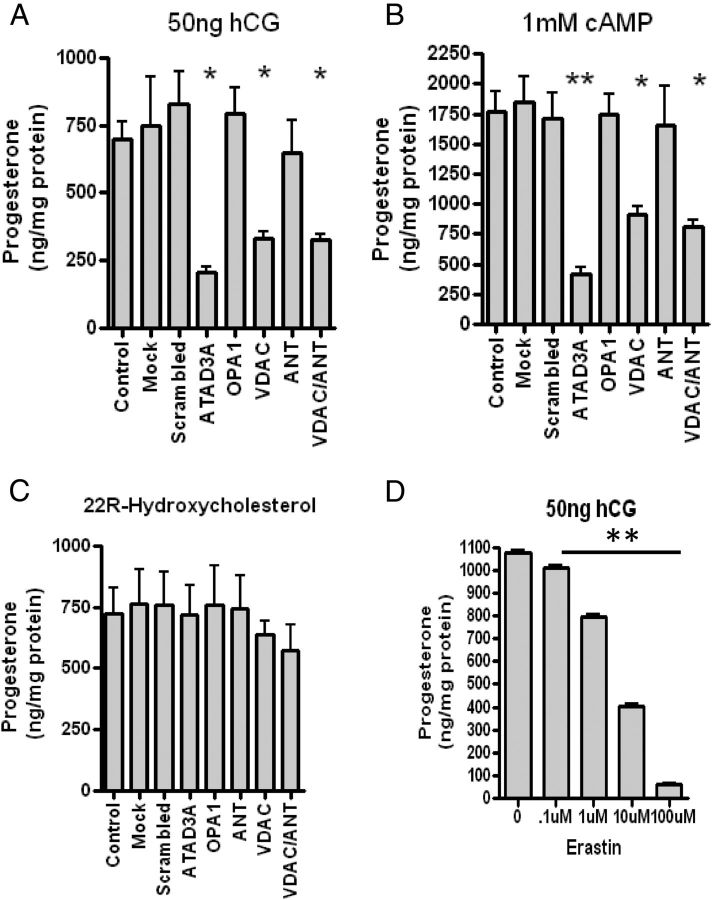
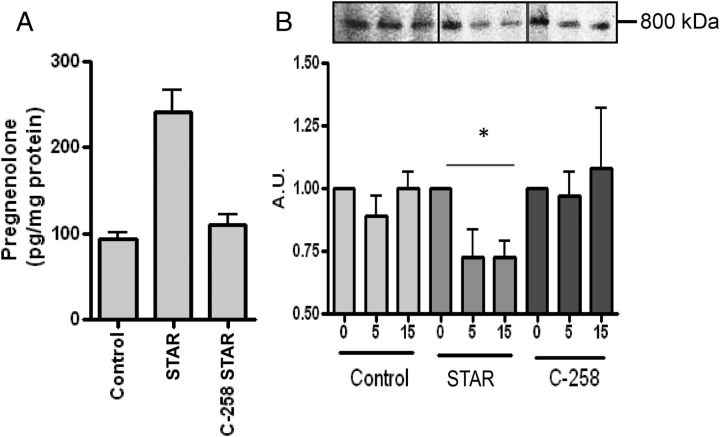
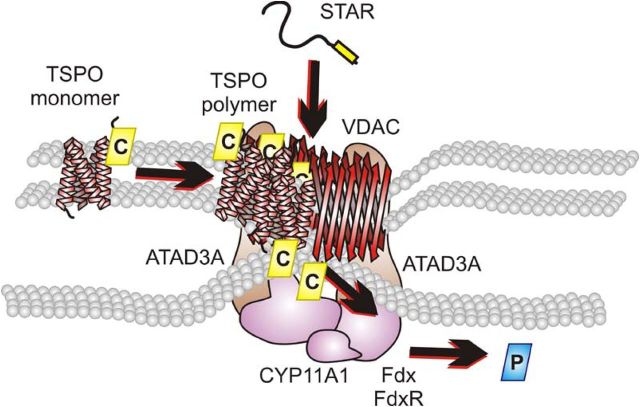
Steroid hormones are critical for organismal development and health. The rate-limiting step in steroidogenesis is the transport of cholesterol from the outer mitochondrial membrane (OMM) to the cytochrome P450 enzyme CYP11A1 in the inner mitochondrial membrane (IMM). Cholesterol transfer occurs through a complex termed the "transduceosome," in which cytosolic steroidogenic acute regulatory protein interacts with OMM proteins translocator protein and voltage-dependent anion channel (VDAC) to assist with the transfer of cholesterol to OMM. It has been proposed that cholesterol transfer from OMM to IMM occurs at specialized contact sites bridging the two membranes composed of VDAC and IMM adenine nucleotide translocase (ANT). Blue native PAGE of Leydig cell mitochondria identified two protein complexes that were able to bind cholesterol at 66- and 800-kDa. Immunoblot and mass spectrometry analyses revealed that the 800-kDa complex contained the OMM translocator protein (18-kDa) and VDAC along with IMM CYP11A1, ATPase family AAA domain-containing protein 3A (ATAD3A), and optic atrophy type 1 proteins, but not ANT. Knockdown of ATAD3A, but not ANT or optic atrophy type 1, in Leydig cells resulted in a significant decrease in hormone-induced, but not 22R-hydroxycholesterol-supported, steroid production. Using a 22-phenoxazonoxy-5-cholene-3-beta-ol CYP11A1-specific probe, we further demonstrated that the 800-kDa complex offers the microenvironment needed for CYP11A1 activity. Addition of steroidogenic acute regulatory protein to the complex mobilized the cholesterol bound at the 800-kDa complex, leading to increased steroid formation. These results identify a bioactive, multimeric protein complex spanning the OMM and IMM unit that is responsible for the hormone-induced import, segregation, targeting, and metabolism of cholesterol.
Figures
References
-
- Srere PA. 1987. Complexes of sequential metabolic enzymes. Annu Rev Biochem 56:89–124 - PubMed
-
- Liu J , Rone MB , Papadopoulos V. 2006. Protein-protein interactions mediate mitochondrial cholesterol transport and steroid biosynthesis. J Biol Chem 281:38879–38893 - PubMed
-
- Hauet T , Yao ZX , Bose HS , Wall CT , Han Z , Li W , Hales DB , Miller WL , Culty M , Papadopoulos V. 2005. Peripheral-type benzodiazepine receptor-mediated action of steroidogenic acute regulatory protein on cholesterol entry into Leydig cell mitochondria. Mol Endocrinol 19:540–554 - PubMed
-
- Bose HS , Lingappa VR , Miller WL. 2002. Rapid regulation of steroidogenesis by mitochondrial protein import. Nature 417:87–91 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
