

Cancer in light of experimental evolution
- PMID: 22975007
- PMCID: PMC3457634
- DOI: 10.1016/j.cub.2012.06.065
Cancer in light of experimental evolution
Abstract
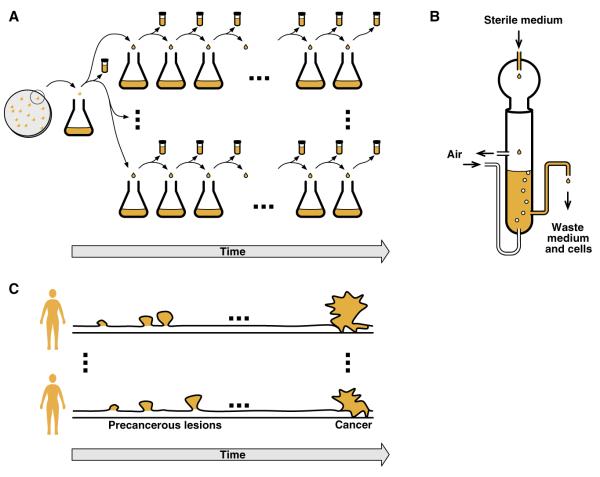
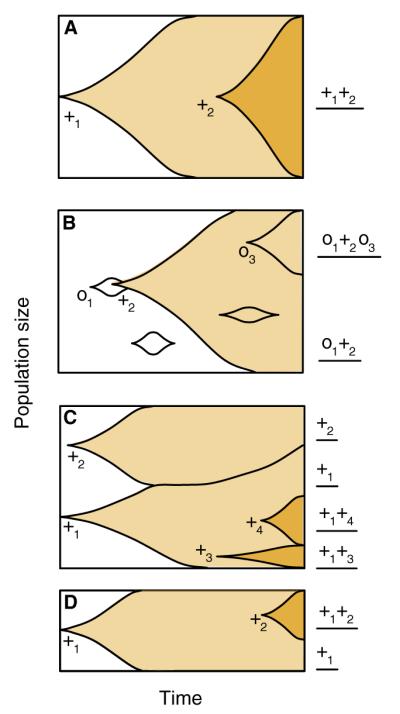
Cancer initiation, progression, and the emergence of therapeutic resistance are evolutionary phenomena of clonal somatic cell populations. Studies in microbial experimental evolution and the theoretical work inspired by such studies are yielding deep insights into the evolutionary dynamics of clonal populations, yet there has been little explicit consideration of the relevance of this rapidly growing field to cancer biology. Here, we examine how the understanding of mutation, selection, and spatial structure in clonal populations that is emerging from experimental evolution may be applicable to cancer. Along the way, we discuss some significant ways in which cancer differs from the model systems used in experimental evolution. Despite these differences, we argue that enhanced prediction and control of cancer may be possible using ideas developed in the context of experimental evolution, and we point out some prospects for future research at the interface between these traditionally separate areas.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Nowell P. The clonal evolution of tumor cell populations. Science. 1976;194:23–28. - PubMed
-
- Merlo L, Pepper J, Reid B, Maley C. Cancer as an evolutionary and ecological process. Nat. Rev. Cancer. 2006;6:924–935. - PubMed
-
- Crespi B, Summers K. Evolutionary biology of cancer. Trends Ecol. Evol. 2005;20:545–552. - PubMed
-
- Frank SA. Dynamics of Cancer: Incidence, Inheritance, and Evolution. Princeton University Press; Princeton: 2007. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
