

TGF-β-miR-34a-CCL22 signaling-induced Treg cell recruitment promotes venous metastases of HBV-positive hepatocellular carcinoma
- PMID: 22975373
- PMCID: PMC3443566
- DOI: 10.1016/j.ccr.2012.07.023
TGF-β-miR-34a-CCL22 signaling-induced Treg cell recruitment promotes venous metastases of HBV-positive hepatocellular carcinoma
Abstract
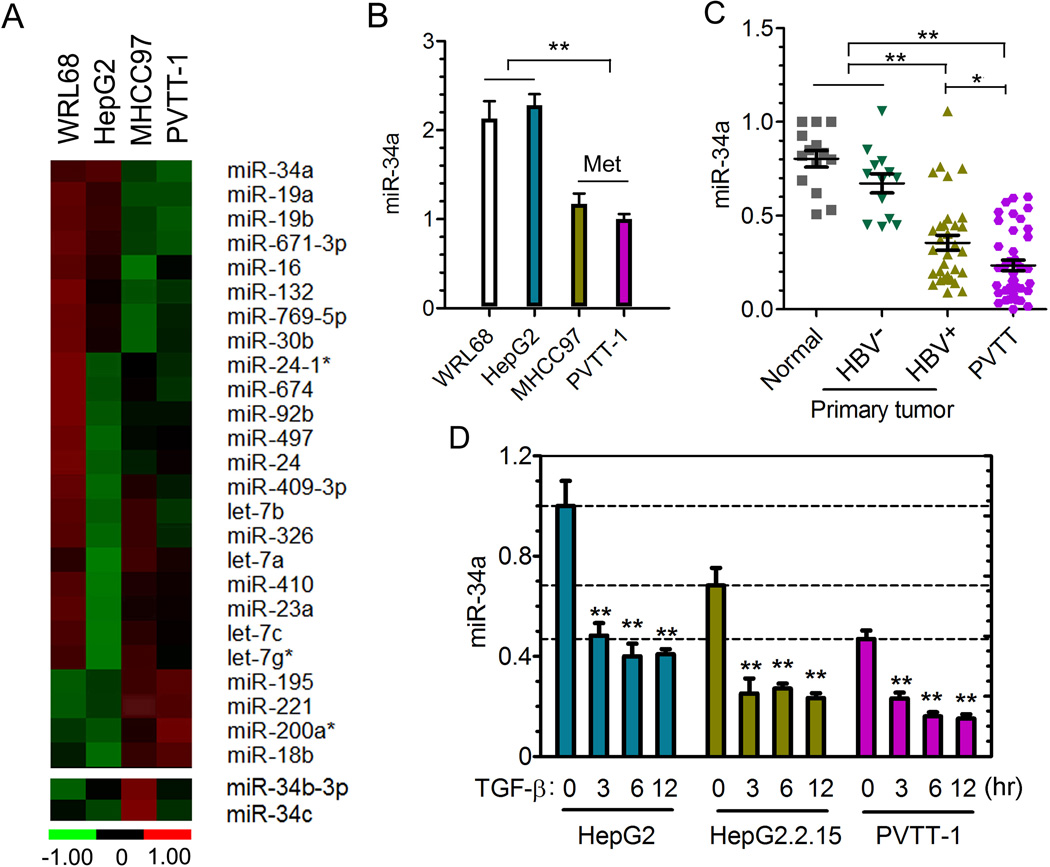
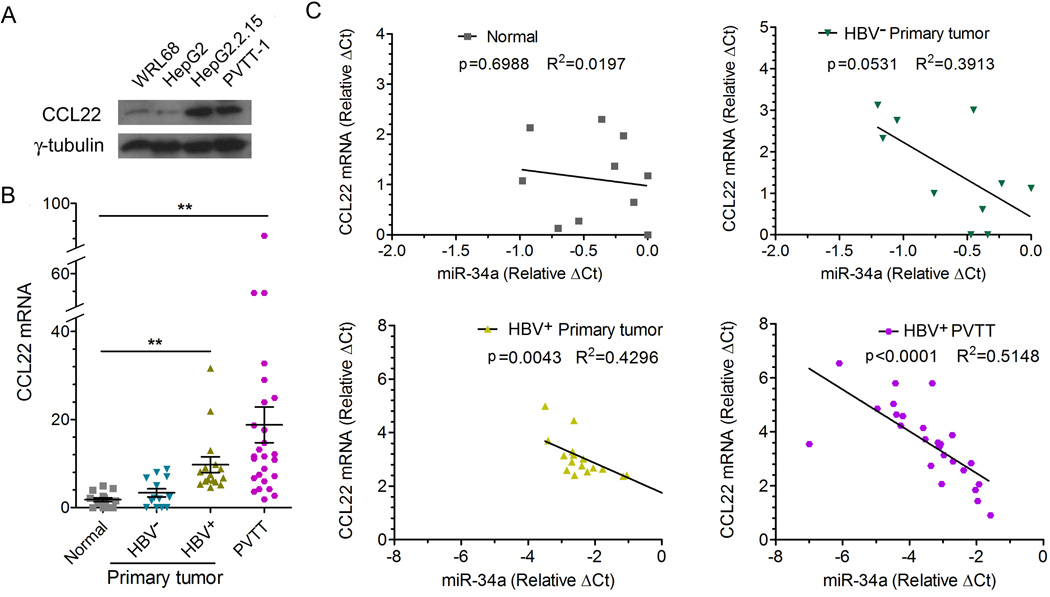
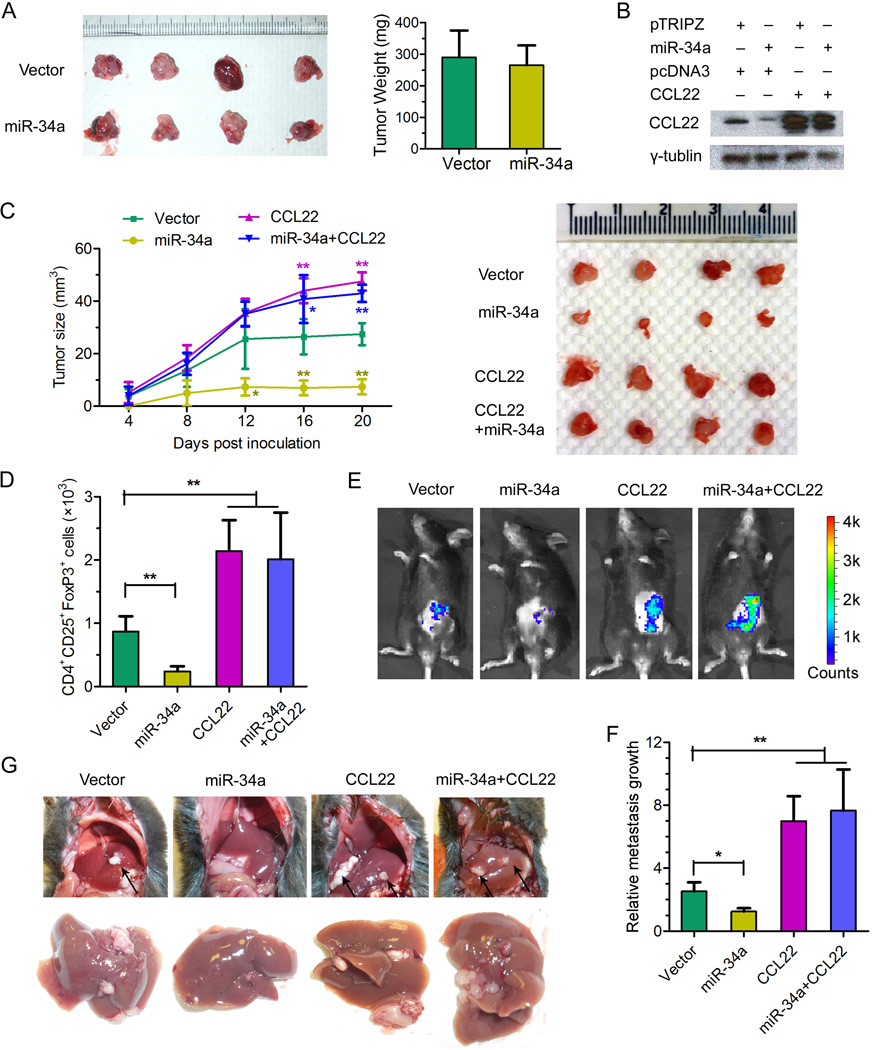
Portal vein tumor thrombus (PVTT) is strongly correlated to a poor prognosis for patients with hepatocellular carcinoma (HCC). In this study, we uncovered a causative link between hepatitis B virus (HBV) infection and development of PVTT. Mechanistically, elevated TGF-β activity, associated with the persistent presence of HBV in the liver tissue, suppresses the expression of microRNA-34a, leading to enhanced production of chemokine CCL22, which recruits regulatory T (Treg) cells to facilitate immune escape. These findings strongly suggest that HBV infection and activity of the TGF-β-miR-34a-CCL22 axis serve as potent etiological factors to predispose HCC patients for the development of PVTT, possibly through the creation of an immune-subversive microenvironment to favor colonization of disseminated HCC cells in the portal venous system.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures




Comment in
-
Transforming the microenvironment: a trick of the metastatic cancer cell.Cancer Cell. 2012 Sep 11;22(3):279-80. doi: 10.1016/j.ccr.2012.08.018. Cancer Cell. 2012. PMID: 22975368
References
-
- Bao S, Ouyang G, Bai X, Huang Z, Ma C, Liu M, Shao R, Anderson RM, Rich JN, Wang XF. Periostin potently promotes metastatic growth of colon cancer by augmenting cell survival via the Akt/PKB pathway. Cancer Cell. 2004;5:329–339. - PubMed
-
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Benhenda S, Cougot D, Buendia MA, Neuveut C. Hepatitis B virus X protein molecular functions and its role in virus life cycle and pathogenesis. Advances in cancer research. 2009;103:75–109. - PubMed
-
- Bierie B, Moses HL. Tumour microenvironment: TGFbeta: the molecular Jekyll and Hyde of cancer. Nat Rev Cancer. 2006;6:506–520. - PubMed
-
- Bissell DM. Chronic liver injury, TGF-beta, and cancer. Exp Mol Med. 2001;33:179–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
