

Facilitation and restoration of cognitive function in primate prefrontal cortex by a neuroprosthesis that utilizes minicolumn-specific neural firing
- PMID: 22976769
- PMCID: PMC3505670
- DOI: 10.1088/1741-2560/9/5/056012
Facilitation and restoration of cognitive function in primate prefrontal cortex by a neuroprosthesis that utilizes minicolumn-specific neural firing
Abstract
Objective: Maintenance of cognitive control is a major concern for many human disease conditions; therefore, a major goal of human neuroprosthetics is to facilitate and/or recover the cognitive function when such circumstances impair appropriate decision making.
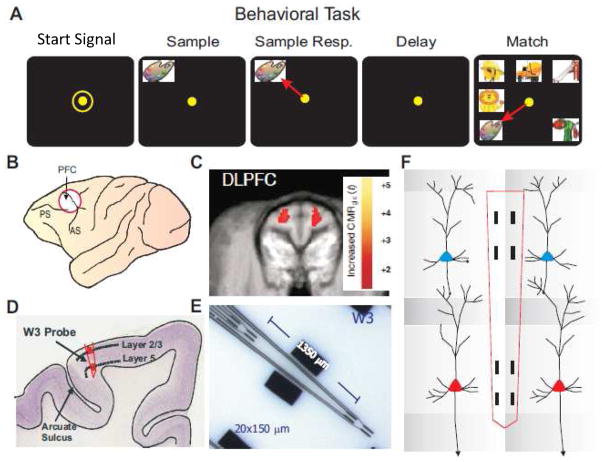
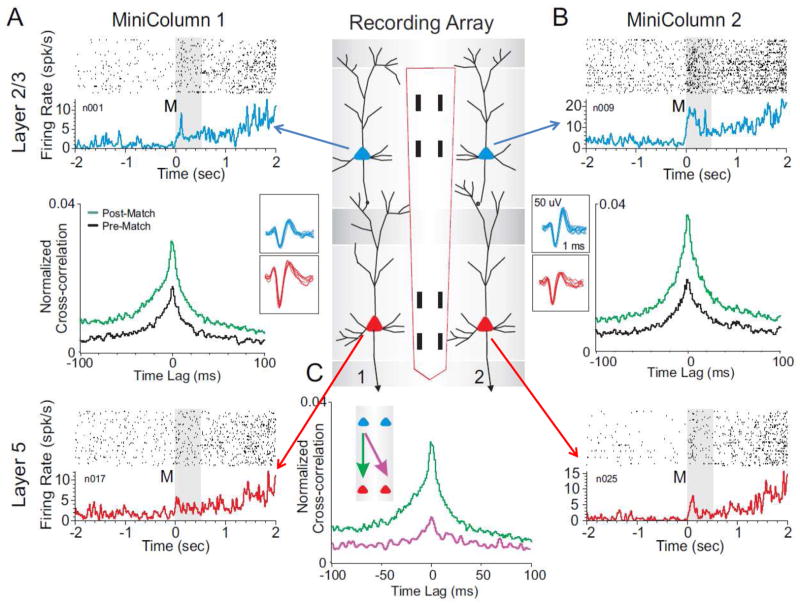
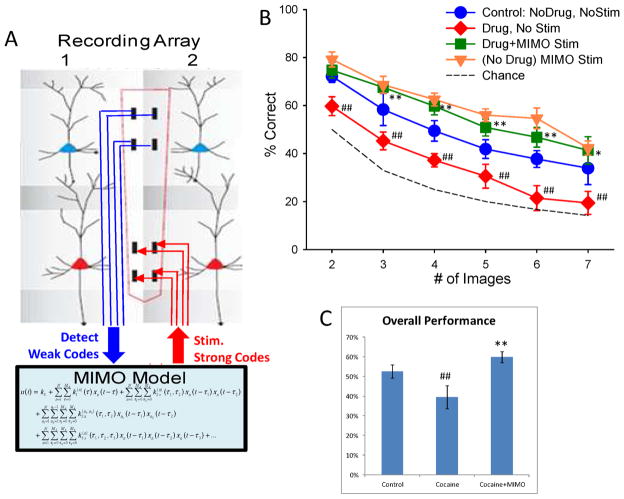
Approach: Minicolumnar activity from the prefrontal cortex (PFC) was recorded from nonhuman primates trained to perform a delayed match to sample (DMS), via custom-designed conformal multielectrode arrays that provided inter-laminar recordings from neurons in the PFC layer 2/3 and layer 5. Such recordings were analyzed via a previously demonstrated nonlinear multi-input-multi-output (MIMO) neuroprosthesis in rodents, which extracted and characterized multicolumnar firing patterns during DMS performance.
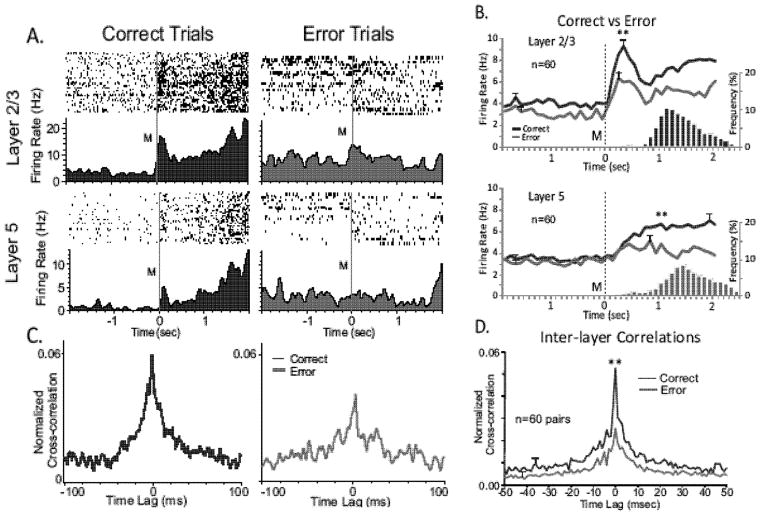
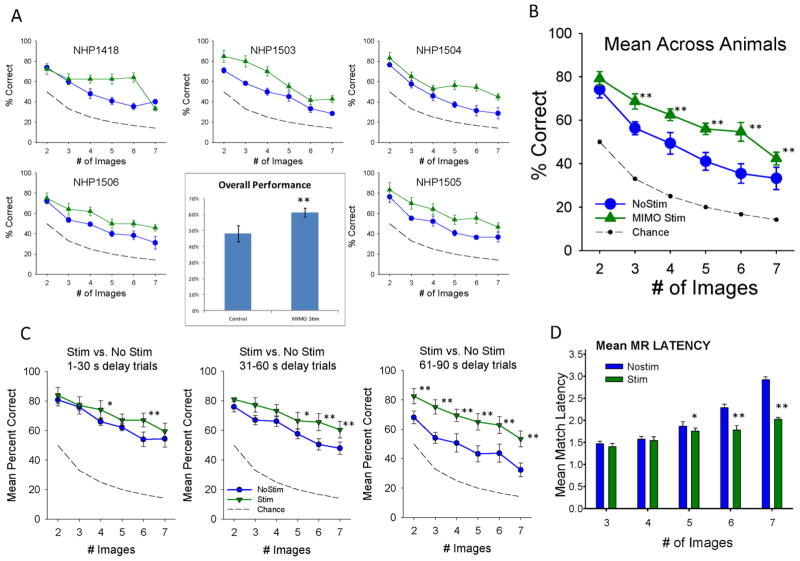
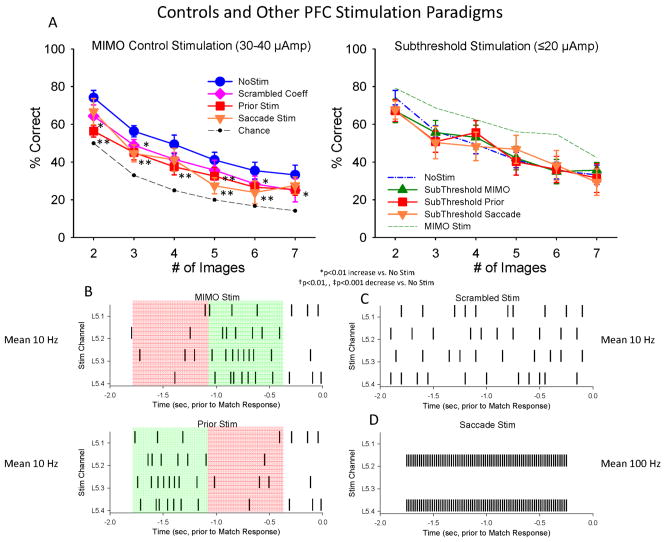
Main results: The MIMO model verified that the conformal recorded individual PFC minicolumns responded to entrained target selections in patterns critical for successful DMS performance. This allowed the substitution of task-related layer 5 neuron firing patterns with electrical stimulation in the same recording regions during columnar transmission from layer 2/3 at the time of target selection. Such stimulation improved normal task performance, but more importantly, recovered performance when applied as a neuroprosthesis following the pharmacological disruption of decision making in the same task.
Significance: These findings provide the first successful application of neuroprosthesis in the primate brain designed specifically to restore or repair the disrupted cognitive function.
Conflict of interest statement
The authors declare no competing financial interests.
Figures


References
-
- Arnsten AF. Stress impairs prefrontal cortical function in rats and monkeys: role of dopamine D1 and norepinephrine alpha-1 receptor mechanisms. Prog Brain Res. 2000;126:183–92. - PubMed
-
- Baddeley A. Fractionating the central executive. In: Stuss DT, Knight RT, editors. Principles of Frontal Lobe Function. New York: Oxford University Press; 2002. pp. 246–260.
-
- Berger TW, Ahuja A, Courellis SH, Deadwyler SA, Erinjippurath G, Gerhardt GA, Gholmieh G, Granacki JJ, Hampson R, Hsaio MC, LaCoss J, Marmarelis VZ, Nasiatka P, Srinivasan V, Song D, Tanguay AR, Wills J. Restoring lost cognitive function. IEEE Eng Med Biol Mag. 2005;24:30–44. - PubMed
-
- Berger TW, Song D, Chan RHM, Marmarelis VZ, LaCoss J, Wills J, Hampson RE, Deadwyler SA, Granacki JJ. A hippocampal cognitive prosthesis: Multi-input, multi-output nonllinear modeling and VLSI instrumentation. IEEE Transactions on Neural Systems and Rehabilitation Engineering. 2012;20:198–211. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous