

What do molecules do when we are not looking? State sequence analysis for stochastic chemical systems
- PMID: 22977098
- PMCID: PMC3481601
- DOI: 10.1098/rsif.2012.0633
What do molecules do when we are not looking? State sequence analysis for stochastic chemical systems
Abstract
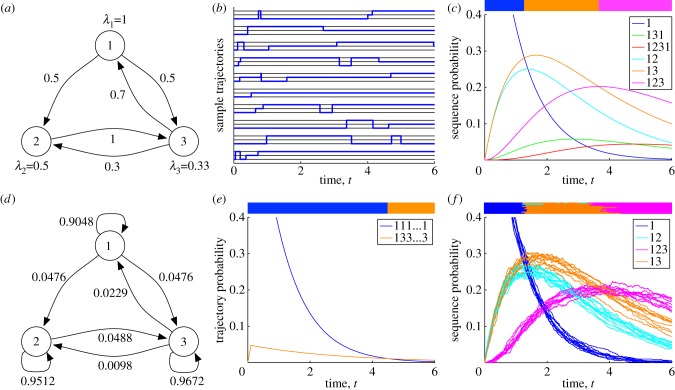
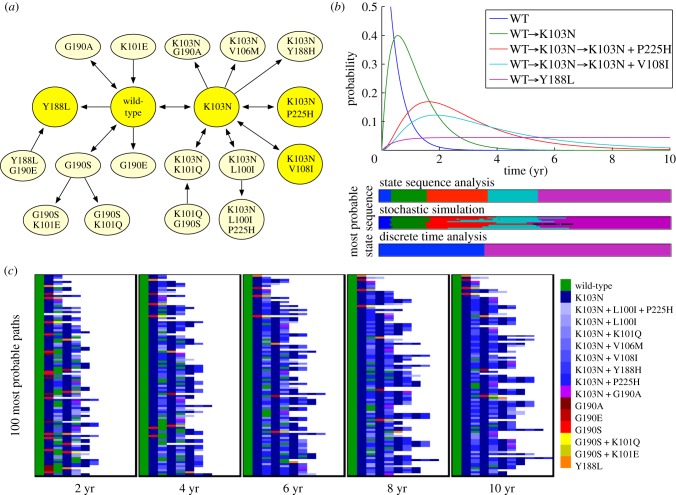
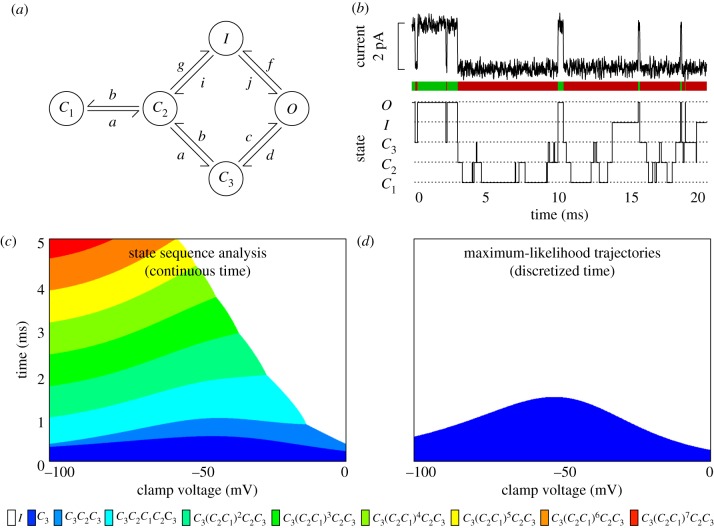
Many biomolecular systems depend on orderly sequences of chemical transformations or reactions. Yet, the dynamics of single molecules or small-copy-number molecular systems are significantly stochastic. Here, we propose state sequence analysis--a new approach for predicting or visualizing the behaviour of stochastic molecular systems by computing maximum probability state sequences, based on initial conditions or boundary conditions. We demonstrate this approach by analysing the acquisition of drug-resistance mutations in the human immunodeficiency virus genome, which depends on rare events occurring on the time scale of years, and the stochastic opening and closing behaviour of a single sodium ion channel, which occurs on the time scale of milliseconds. In both cases, we find that our approach yields novel insights into the stochastic dynamical behaviour of these systems, including insights that are not correctly reproduced in standard time-discretization approaches to trajectory analysis.
Figures
References
-
- Nie S., Chiu D. T., Zare R. N. 1995. Real-time detection of single molecules in solution by confocal fluorescence microscopy. Anal. Chem. 67, 2849–285710.1021/ac00113a019 (doi:10.1021/ac00113a019) - DOI - DOI
-
- Sekar R. B., Periasamy A. 2003. Fluorescence resonance energy transfer (FRET) microscopy imaging of live cell protein localizations. J. Cell Biol. 160, 629–63310.1083/jcb.200210140 (doi:10.1083/jcb.200210140) - DOI - DOI - PMC - PubMed
-
- Swain P. S., Elowitz M. B., Siggia E. D. 2002. Intrinsic and extrinsic contributions to stochasticity in gene expression. Proc. Natl Acad. Sci. USA 99, 12 795–12 80010.1073/pnas.162041399 (doi:10.1073/pnas.162041399) - DOI - DOI - PMC - PubMed
-
- Kepler T. B., Elston T. C. 2001. Stochasticity in transcriptional regulation: origins, consequences, and mathematical representations. Biophys. J. 81, 3116–313610.1016/S0006-3495(01)75949-8 (doi:10.1016/S0006-3495(01)75949-8) - DOI - DOI - PMC - PubMed
-
- Rao C. V., Wolf D. M., Arkin A. P. 2002. Control, exploitation and tolerance of intracellular noise. Nature 420, 231–23710.1038/nature01258 (doi:10.1038/nature01258) - DOI - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
