

Human Tribbles 3 protects nuclear DNA from cytidine deamination by APOBEC3A
- PMID: 22977230
- PMCID: PMC3493958
- DOI: 10.1074/jbc.M112.372722
Human Tribbles 3 protects nuclear DNA from cytidine deamination by APOBEC3A
Abstract
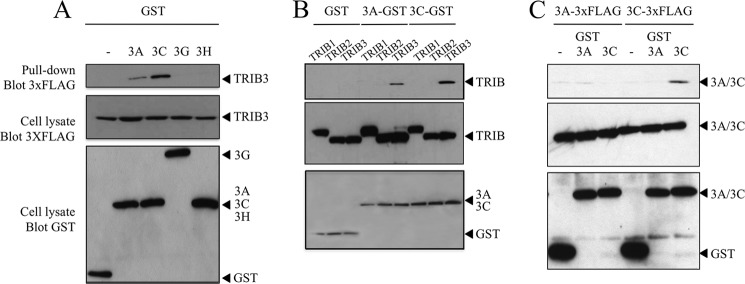
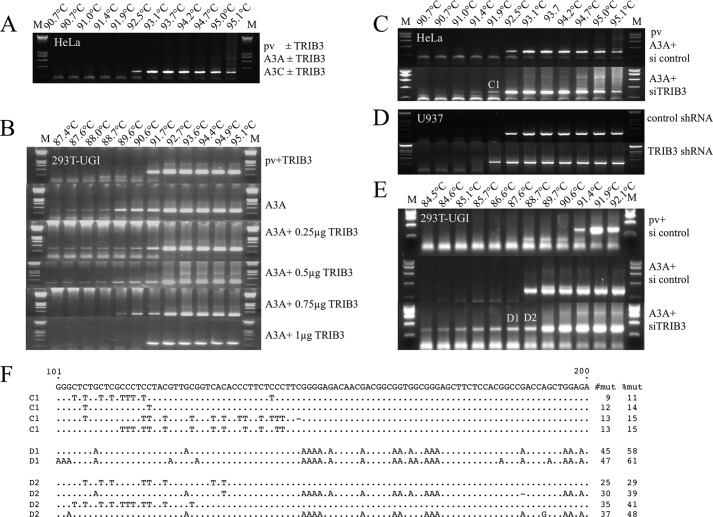
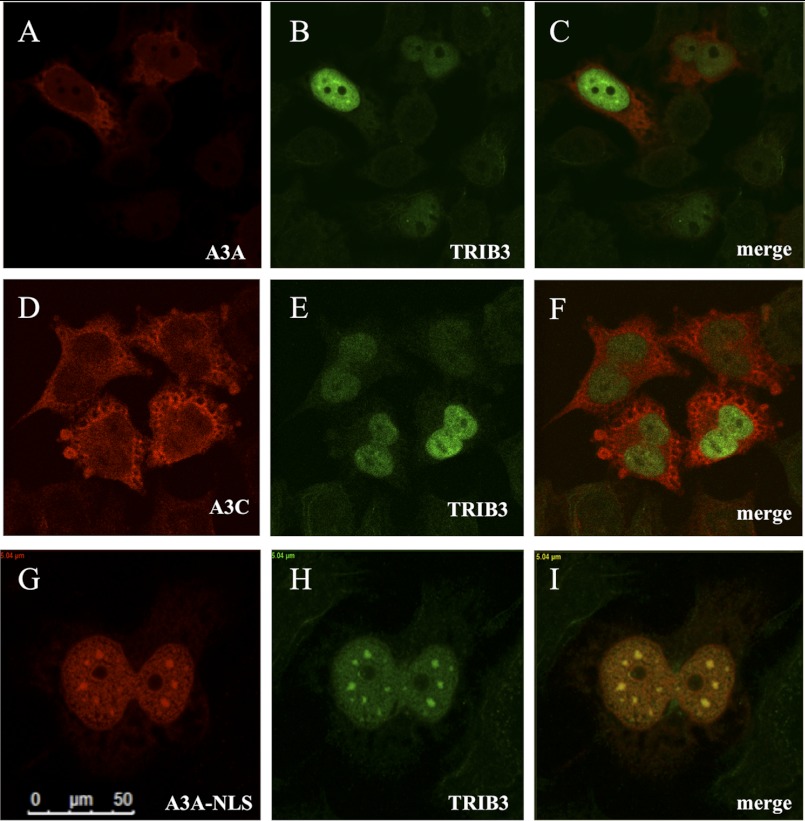
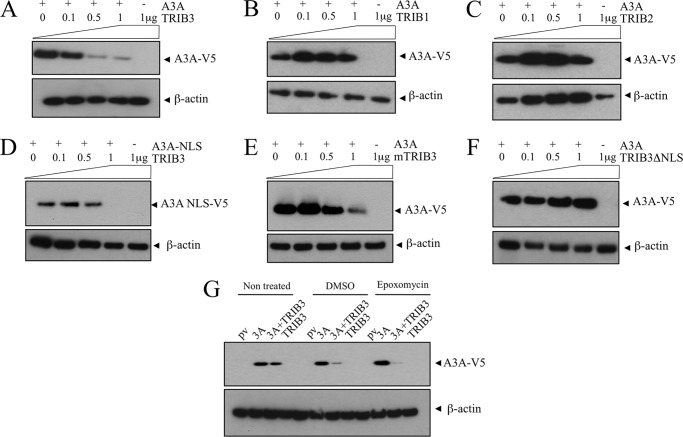
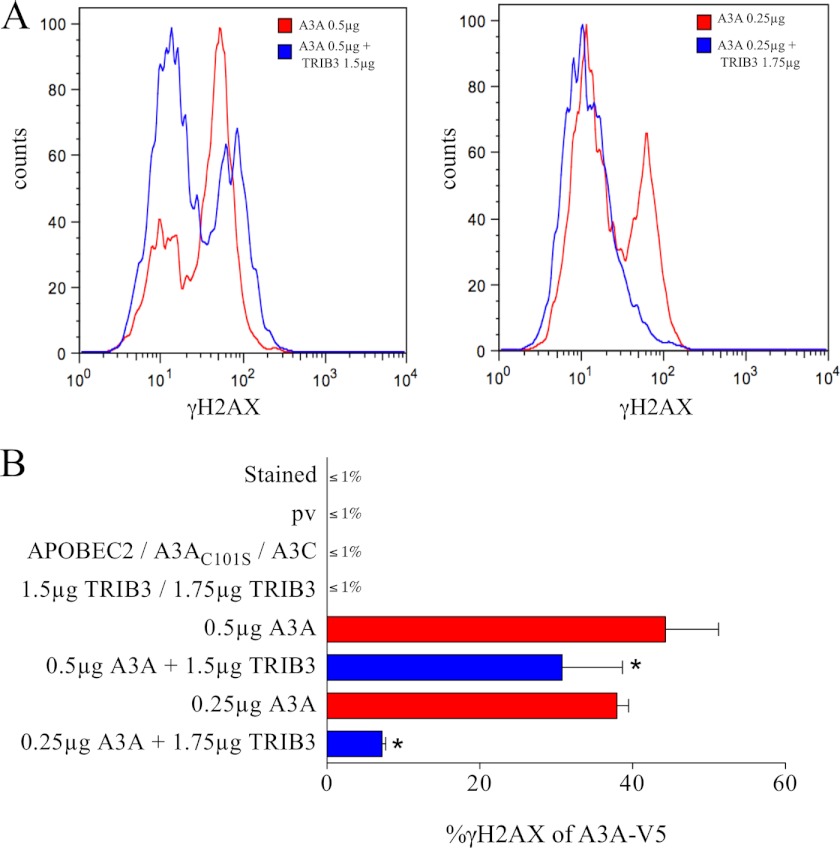
The human polydeoxynucleotide cytidine deaminases APOBEC3A, APOBEC3C, and APOBEC3H are capable of mutating viral DNA in the nucleus, whereas APOBEC3A alone efficiently edits nuclear DNA. Deamination is rapidly followed by excision of uracil residues and can lead to double-stranded breaks. It is not known to which protein networks these DNA mutators belong. Using a yeast two-hybrid screen, we identified the human homolog of Drosophila Tribbles 3, TRIB3, as an interactor for APOBEC3A and APOBEC3C. The interaction was confirmed by co-affinity purification. Co-transfection of APOBEC3A with a TRIB3 expression vector reduced nuclear DNA editing whereas siRNA knockdown of TRIB3 increased the levels of nuclear DNA editing, indicating that TRIB3 functioned as a repressor of A3A. It also repressed A3A-associated γH2AX positive double-stranded breaks. The interaction results in degradation of A3A in a proteasome-independent manner. TRIB3 has been linked to cancer and via its own interactors and links the A3A DNA mutators to the Rb-BRCA1-ATM network. TRIB3 emerges as an important guardian of genome integrity.
Figures

Similar articles
-
Efficient deamination of 5-methylcytidine and 5-substituted cytidine residues in DNA by human APOBEC3A cytidine deaminase.PLoS One. 2013 Jun 20;8(6):e63461. doi: 10.1371/journal.pone.0063461. Print 2013. PLoS One. 2013. PMID: 23840298 Free PMC article.
-
The cytidine deaminase APOBEC3C has unique sequence and genome feature preferences.Genetics. 2024 Aug 7;227(4):iyae092. doi: 10.1093/genetics/iyae092. Genetics. 2024. PMID: 38946641 Free PMC article.
-
Orthologous mammalian APOBEC3A cytidine deaminases hypermutate nuclear DNA.Mol Biol Evol. 2014 Feb;31(2):330-40. doi: 10.1093/molbev/mst195. Epub 2013 Oct 25. Mol Biol Evol. 2014. PMID: 24162735
-
Advantages and disadvantages of cytidine deamination.J Immunol. 2004 Jun 1;172(11):6513-8. doi: 10.4049/jimmunol.172.11.6513. J Immunol. 2004. PMID: 15153462 Review.
-
Generation of Genomic Alteration from Cytidine Deamination.Adv Exp Med Biol. 2018;1044:49-64. doi: 10.1007/978-981-13-0593-1_5. Adv Exp Med Biol. 2018. PMID: 29956291 Review.
Cited by
-
APOBEC3H Subcellular Localization Determinants Define Zipcode for Targeting HIV-1 for Restriction.Mol Cell Biol. 2018 Nov 13;38(23):e00356-18. doi: 10.1128/MCB.00356-18. Print 2018 Dec 1. Mol Cell Biol. 2018. PMID: 30224517 Free PMC article.
-
Specific and nonhepatotoxic degradation of nuclear hepatitis B virus cccDNA.Science. 2014 Mar 14;343(6176):1221-8. doi: 10.1126/science.1243462. Epub 2014 Feb 20. Science. 2014. PMID: 24557838 Free PMC article.
-
Genotoxic stress increases cytoplasmic mitochondrial DNA editing by human APOBEC3 mutator enzymes at a single cell level.Sci Rep. 2019 Feb 28;9(1):3109. doi: 10.1038/s41598-019-39245-8. Sci Rep. 2019. PMID: 30816165 Free PMC article.
-
Molecular basis of the attenuated phenotype of human APOBEC3B DNA mutator enzyme.Nucleic Acids Res. 2015 Oct 30;43(19):9340-9. doi: 10.1093/nar/gkv935. Epub 2015 Sep 17. Nucleic Acids Res. 2015. PMID: 26384561 Free PMC article.
-
An overview of the functions and mechanisms of APOBEC3A in tumorigenesis.Acta Pharm Sin B. 2024 Nov;14(11):4637-4648. doi: 10.1016/j.apsb.2024.08.020. Epub 2024 Aug 27. Acta Pharm Sin B. 2024. PMID: 39664421 Free PMC article. Review.
References
-
- Di Noia J. M., Neuberger M. S. (2007) Molecular mechanisms of antibody somatic hypermutation. Annu. Rev. Biochem. 76, 1–22 - PubMed
-
- Jarmuz A., Chester A., Bayliss J., Gisbourne J., Dunham I., Scott J., Navaratnam N. (2002) An anthropoid-specific locus of orphan C to U RNA-editing enzymes on chromosome 22. Genomics 79, 285–296 - PubMed
-
- Harris R. S., Bishop K. N., Sheehy A. M., Craig H. M., Petersen-Mahrt S. K., Watt I. N., Neuberger M. S., Malim M. H. (2003) DNA deamination mediates innate immunity to retroviral infection. Cell 113, 803–809 - PubMed
-
- Lecossier D., Bouchonnet F., Clavel F., Hance A. J. (2003) Hypermutation of HIV-1 DNA in the absence of the Vif protein. Science 300, 1112. - PubMed
-
- Mangeat B., Turelli P., Caron G., Friedli M., Perrin L., Trono D. (2003) Broad antiretroviral defense by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature 424, 99–103 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
