

Mono- and dithiol glutaredoxins in the trypanothione-based redox metabolism of pathogenic trypanosomes
- PMID: 22978520
- PMCID: PMC3739957
- DOI: 10.1089/ars.2012.4932
Mono- and dithiol glutaredoxins in the trypanothione-based redox metabolism of pathogenic trypanosomes
Abstract
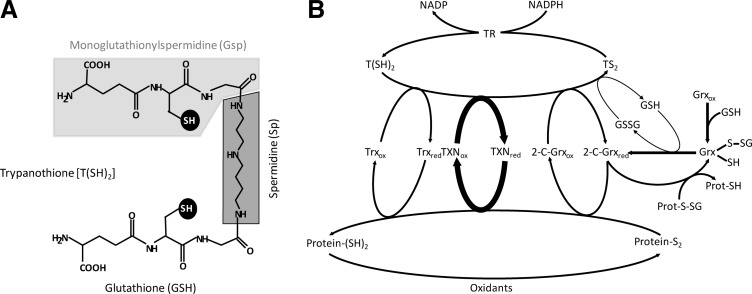
Significance: Glutaredoxins are ubiquitous small thiol proteins of the thioredoxin-fold superfamily. Two major groups are distinguished based on their active sites: the dithiol (2-C-Grxs) and the monothiol (1-C-Grxs) glutaredoxins with a CXXC and a CXXS active site motif, respectively. Glutaredoxins are involved in cellular redox and/or iron sulfur metabolism. Usually their functions are closely linked to the glutathione system. Trypanosomatids, the causative agents of several tropical diseases, rely on trypanothione as principal low molecular mass thiol, and their glutaredoxins readily react with the unique bis(glutathionyl) spermidine conjugate.
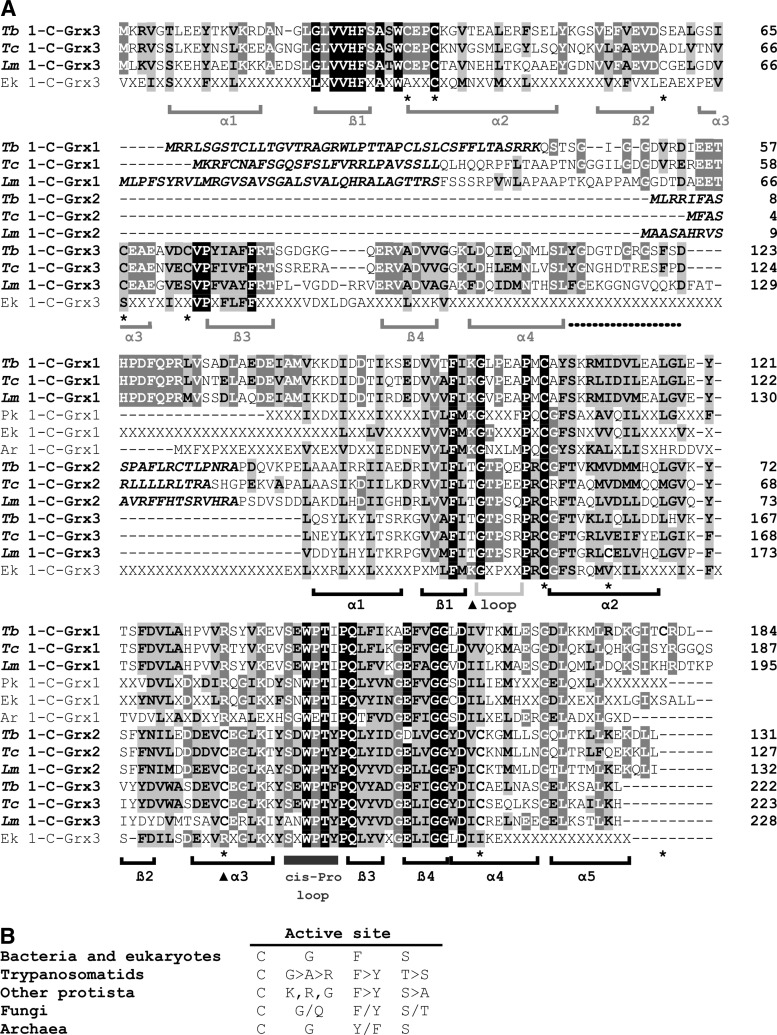
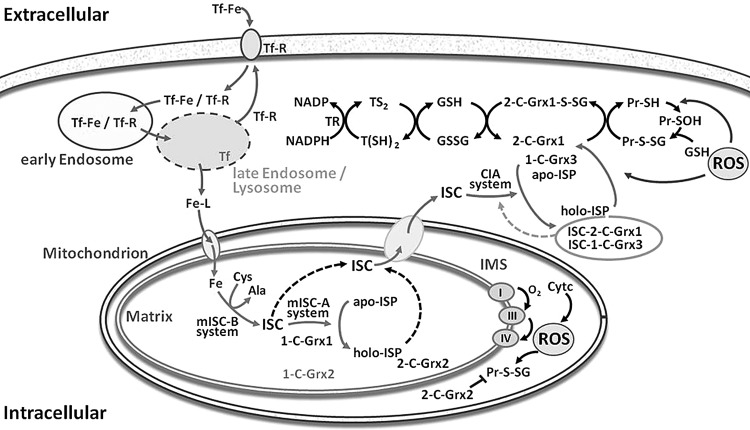
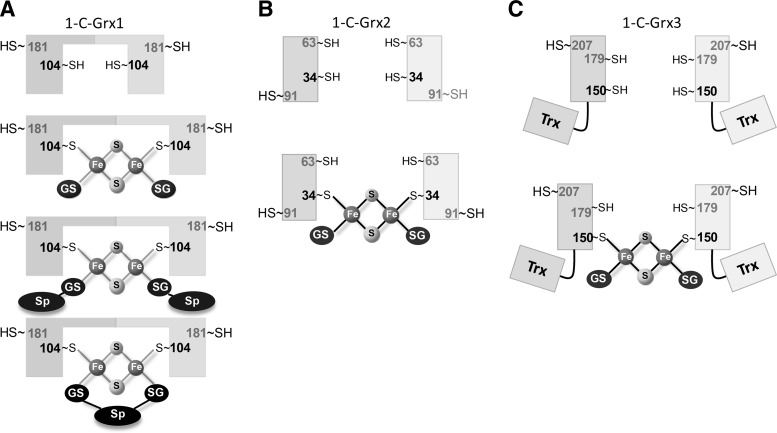
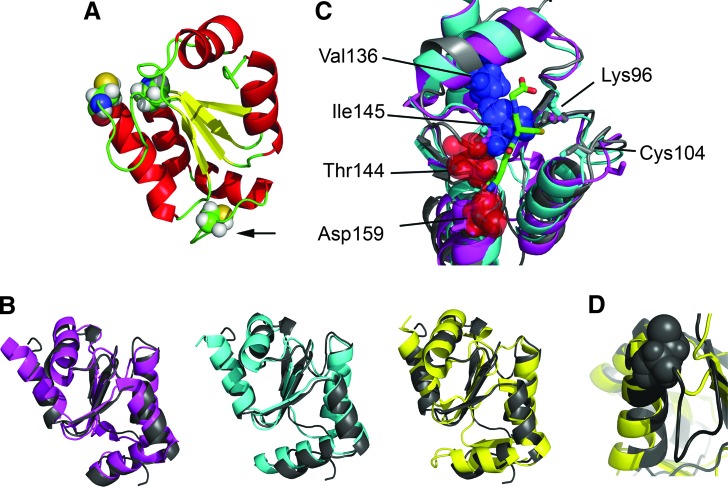
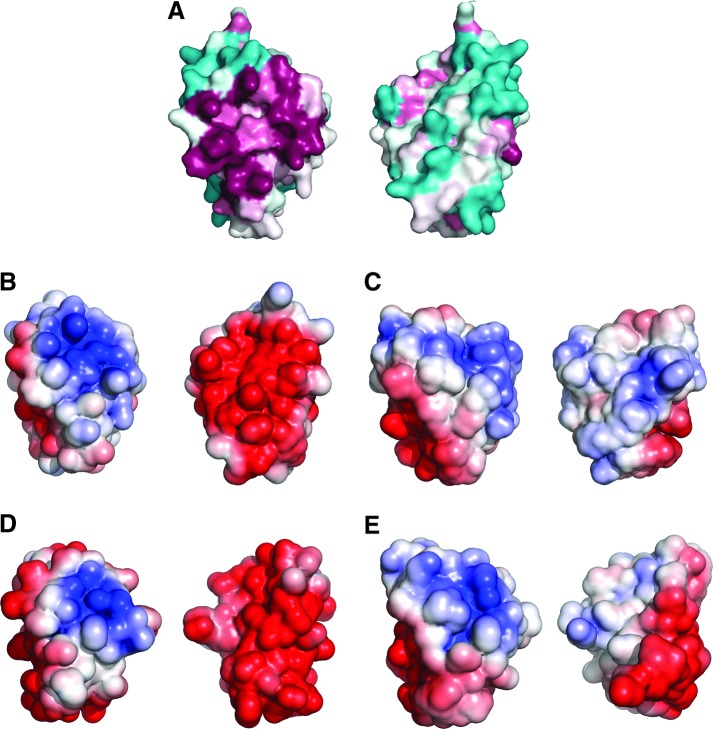
Recent advances: Two 2-C-Grxs and three 1-C-Grxs have been identified in pathogenic trypanosomatids. The 2-C-Grxs catalyze the reduction of glutathione disulfide by trypanothione and display reductase activity towards protein disulfides, as well as protein-glutathione mixed disulfides. In vitro, all three 1-C-Grxs as well as the cytosolic 2-C-Grx of Trypanosoma brucei can complex an iron-sulfur cluster. Recently the structure of the 1-C-Grx1 has been solved by NMR spectroscopy. The structure is very similar to those of other 1-C-Grxs, with some differences in the loop containing the conserved cis-Pro and the surface charge distribution.
Critical issues: Although four of the five trypanosomal glutaredoxins proved to coordinate an iron-sulfur cluster in vitro, the physiological role of the mitochondrial and cytosolic proteins, respectively, has only started to be unraveled.
Future directions: The use of trypanothione by the glutaredoxins has established a novel role for this parasite-specific dithiol. Future work should reveal if these differences can be exploited for the development of novel antiparasitic drugs.
Figures
References
-
- Alphey MS. Gabrielsen M. Micossi E. Leonard GA. McSweeney SM. Ravelli RB. Tetaud E. Fairlamb AH. Bond CS. Hunter WN. Tryparedoxins from Crithidia fasciculata and Trypanosoma brucei: photoreduction of the redox disulfide using synchrotron radiation and evidence for a conformational switch implicated in function. J Biol Chem. 2003;278:25919–25925. - PubMed
-
- Arias DG. Cabeza MS. Erben ED. Carranza PG. Lujan HD. Tellez Inon MT. Iglesias AA. Guerrero SA. Functional characterization of methionine sulfoxide reductase A from Trypanosoma spp. Free Radic Biol Med. 2011;50:37–46. - PubMed
-
- Askelöf P. Axelsson K. Eriksson S. Mannervik B. Mechanism of action of enzymes catalyzing thiol-disulfide interchange. Thioltransferases rather than transhydrogenases. FEBS Lett. 1974;38:263–267. - PubMed
-
- Beer SM. Taylor ER. Brown SE. Dahm CC. Costa NJ. Runswick MJ. Murphy MP. Glutaredoxin 2 catalyzes the reversible oxidation and glutathionylation of mitochondrial membrane thiol proteins: Implications for mitochondrial redox regulation and antioxidant defense. J Biol Chem. 2004;279:47939–47951. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
